Бластогенез ланцетника. Дробление яйцеклетки ланцетника
Обновлено: 18.04.2024
1. Антипчук, Ю.П. Гистология с основами эмбриологии / Ю.П. Антипчук. – М.: Просвещение, 1983. – 240 с.
2. Алмазов, И.В., Сутулов Л.С. Атлас по гистологии и эмбриологии / И.В. Алмазов, Л.С. Сутулов. – М.: Медицина, 1978. – 148 с.
3. Гистология / под ред. Ю.И. Афанасьева. – М: Медицина, 1989. – 361 с.
4. Рябов, К.П. Гистология с основами эмбриологии / К.П. Рябов. – Мн.: Высш. шк., 1991. – 289 с.
5. Биологический энциклопкдический словарь / под ред. М.С. Гилярова. – М.: Сов. Энцикл., 1989. – 864 с.
6. Практикум по гистологии, цитологии и эмбриологии / под ред. Н.А. Юриной, А.И. Радостиной. – М.: Высш. шк., 1989. – 154 с.
Хэм А., Кормик Д. Гистология / А. Хэм, Д. Кормик. – М.: Мир, 1983. – 192
1. Особенности эмбрионального развития млекопитающих.
2. Эмбриогенез яйцекладущих млекопитающих.
3. Эмбриогенез сумчатых млекопитающих.
4. Эмбриогенез плацентарных млекопитающих.
5. Эмбриогенез человека.
Общая характеристика анамний и амниот
Общая характеристика анамний
Исходя из особенностей эмбрионального развития, все хордовые подразделяются на две группы: анамнии и амниоты. Анамнии – это животные, у которых в процессе эмбрионального развития не образуются такие зародышевые оболочки, как амнион, или водная оболочка, и аллантоис. К анамниям относят хордовых, ведущих первичноводный образ жизни, а также низших хордовых, тесно связанных с водной средой в период размножения и эмбрионального развития зародышей – бесчелюстных, рыб и земноводных. В связи с эмбриональным развитием этих хордовых в водной среде, у них отсутствует водная оболочка и аллантоис, так как функции дыхания, выделения и питания развивающегося зародыша обеспечивает окружающая его водная среда.
Хордовых, относящихся к анамниям по характеру эмбрионального развития можно подразделить на три группы:
1) ланцетник, яйцеклетки которого содержит мало желтка;
2) некоторые круглоротые, рыбы (хрящевые ганоиды) и земноводные, яйцеклетки которых содержат среднее количество желтка;
3) селяхии и костистые рыбы, яйцеклетки содержат много желтка.
Эмбриогенез ланцетника
После оплодотворения в яйцеклетке ланцетника начинается перераспределение желтка, который концентрируется в основном на одной стороне яйцеклетки, соответствующей вегетативному полюсу. Анимальный полюс яйцеклетки определяется по расположенному над ним второму полярному тельцу. Дробление яйцеклетки полное, равномерное (рисунок 1).

/ – анимальный полюс; 2 – вегетативный полюс; 3 –скопление желтка; 4 –целобластула; 5 – клетки бластодермы.
Рисунок–1. Последовательность (I–VI) дробления яйцеклетки ланцетника
Первые два дробления идут меридионально, третье – экваториально. Дальнейшее дробление идет попеременно то в одном, то в другом направлении, и количество клеток увеличивается в геометрической прогрессии. После образования однослойного зародыша – бластулы становится заметным, что клетки анимального полюса более мелкие, чем клетки вегетативного полюса. В шарообразной целобластуле ланцетника различают уплощенную часть вегетативного полюса, называемую дном бластулы, а противоположную часть, соответствующую анимальному полюсу, называют крышей бластулы. Клетки, образующие крышу бластулы, будут дифференцироваться в клетки наружного зародышевого листка, или эктодерму, а клетки дна бластулы – в энтодерму.
Гаструляция осуществляется путем инвагинации бластодермы вегетативного полюса внутрь бластоцеля. Впячивание продолжается до тех пор, пока клетки вегетативного полюса соприкоснутся с клетками анимального полюса, в связи с чем полость бластоцеля суживается и исчезает (рисунок 2).
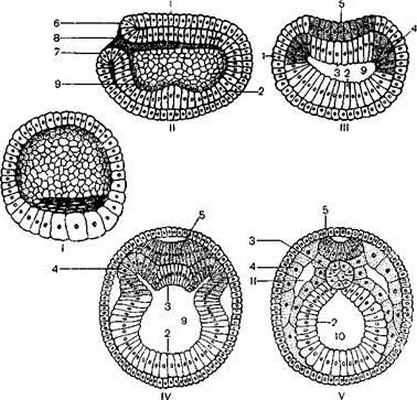
I– целобластула; II – IV – гаструляция; V–нейрула;
1 – эктодерма; 2–энтодерма; 3 –хорда; 4–мезодерма; 5 – нервная пластинка; 6 –верхняя и 7 – нижняя губа бластопора; 8 – бластопор; 9–полость первичной кишки; 10– полость вторичной кишки; 11 – целом.
Рисунок 2–Эмбриогенез ланцетника
С завершением первого этапа гаструляции возникает двуслойный зародыш, или гаструла, состоящий из клеток наружного зародышевого листка – эктодермы и внутреннего зародышевого листка – энтодермы. В результате впячивания образуется полость первичной кишки, выстланная клетками энтодермы, которая сообщается с внешней средой бластопором. Клеточный состав энтодермы неоднородный, поскольку в нее входит также клеточный материал будущей хорды и мезодермы. С образованием полости первичной кишки зародыш начинает быстро расти и удлиняется, но наиболее интенсивные формообразовательные процессы осуществляются в области верхней, или дорсальной, губы бластопора. Непосредственно за верхней губой бластопора, на спинной поверхности зародыша, эктодерма утолщается и состоит из высоких призматических клеток, называемых медуллярной или нервной пластинкой. Эктодерма, окружающая нервную пластинку, представлена мелкими клетками, которые образуют кожный покров. Под нервной пластинкой такие же изменения претерпевают клетки энтодермы, которые представляют материал будущей хорды. В дальнейшем нервная пластинка начинает прогибаться, образуя нервный желобок, а клетки кожной эктодермы интенсивно наползают на него. Впоследствии нервный желобок углубляется, края его смыкаются, и он превращается в нервную трубку, полость которой именуется, нервным каналом. Клетки кожной эктодермы смыкаются, и нервная трубка оказывается под ними. Одновременно клетки энтодермы, примыкающие к нервной пластинке, прогибаются в сторону последней, скручиваются и обособляются в плотный тяж – хорду, которая имеет вид сплошного цилиндра. По сторонам от хордального зачатка энтодерма инвагинирует в сторону эктодермы, образуя мезодермальные выпячивания, или мезодермальные мешки, которые впоследствии отшнуровываются от энтодермы и начинают разрастаться между эктодермой и энтодермой. Полость мезодермальных мешков, возникающая из гастроцеля, превращается во вторичную полость тела, или целом. Таким образом, в процессе гаструляции возникает трехслойный зародыш.
После обособления хорды и отшнуровывания мезодермальных мешков края энтодермы постепенно сближаются в спинной части зародыша и, смыкаясь, образуют замкнутую кишечную трубку. Вслед за гаструляцией у зародыша возникает комплекс осевых органов, характерный для представителей типа хордовых. Он состоит из хорды, по бокам которой располагаются скопления сегментированной мезодермы – сомиты.
Закладка осевых органов происходит на стадии нейрулы. Нервная трубка ланцетника в передней и задней частях зародыша некоторое время остается открытой. В дальнейшем на задней части тела зародыша эктодерма нарастает на бластопор и закрывает его так, что полость нервной трубки сообщается с кишечной полостью нервно-кишечным каналом, который быстро зарастает. Ротовое отверстие у зародыша ланцетника образуется вторично на переднем конце тела вследствие истончения и прорыва эктодермы.
Третий зародышевый листок, или мезодерма, зародыша ланцетника сегментирована на всем протяжении. Мезодермальные сегменты в дальнейшем разделяются на спинную часть – сомиты и брюшную часть – спланхнотомы. Сомиты остаются сегментированными, а спланхнотомы на каждой стороне тела утрачивают первичную сегментацию, сливаются и образуют, расщепляясь на два листка, правую и левую целомические полости. Последние объединяются под кишечной трубкой в общую вторичную полость тела. Когда у ланцетника начинает формироваться хвост, то нервно-кишечный канал исчезает, а на заднем конце зародыша на месте бластопора вследствие истончения и прорыва стенки тела возникает анальное отверстие. Пройдя описанные стадии развития, ланцетник становится свободноплавающей личинкой. В период личиночного развития завершается органогенез и гистогенез и личинка превращается во взрослое животное.
Бластогенез ланцетника. Дробление яйцеклетки ланцетника
Бластогенез ланцетника. Дробление яйцеклетки ланцетника
Ланцетник (Amphioxus lanceolatus, или Branchiostoma lanceolatum) — филогенетически наиболее низко организованный представитель животных (Chordata), у которого в процессе развития уже появляется прочная опора спинного отдела тела — является отличным объектом для схематической демонстрации всех принципиальных бластогенетических процессов, характерных для всего разряда позвоночных животных.
Amphioxus — это маленькое животное, по внешнему виду напоминающее маленькую рыбку; он водится во влажном песке на морских побережьях. Развитие ланцетника было впервые описано русским эмбриологом А. О. Ковалевским.
Выше уже упоминалось о том, какое значение имеет изучение основных процессов развития ланцетника и низших позвоночных для понимания развития человека. В их развитии наблюдаются исторические (филогенетические) факторы и процессы, объяснимые (лишь в том случае, если учитываются филогенетические взаимосвязи развития человека с остальными позвоночными) в общем с точки зрения основного биогенетического закона о схематическом повторении филогенетического развития в онтогенезе. В результате изменений внешней и внутренней жизненной среды, в процессе филогенетического развития к онтогенезу высших животных присоединились ценогенетические факторы, которые включались, ассимилировались и аккумулировались вместе с основными палингенетическими элементами родового развития в живом веществе половых клеток.
В связи с этим без сравнительного изучения, без исследования филогенеза человека, иными словами, без учета хотя бы основных черт развития низших позвоночных невозможно понять полностью сложные процессы развития, происходящие во время онтогенетического развития высших млекопитающих и человека.

Яйцеклетки ланцетника являются олиголецитальными и даже почти изолецитальными, с весьма умеренным превалированием желтка на вегетативном полюсе. Таким образом, они относятся к голобластическому виду, и их дробление проходит по типу полного равномерного, или правильного, дробления. При дроблении, в сущности, происходит митотическое деление, при котором обе вновь возникшие клетки, называемые бластомерами, не расходятся, а остаются вместе, тесно примыкая друг к другу. В плоскости, в которой яйцеклетка делится на два самостоятельных бластомера, на ее поверхности образуется сначала поверхностная, а затем более глубокая борозда, которая постепенно охватывает всю окружность шаровидной клетки.
Эта борозда соответствует цитокинетической перетяжке делящейся митотически клетки в телофазе.
Первая борозда начинает формироваться на анимальном полюсе яйцеклетки сначала в виде малой, поверхностной насечки, которая, непрестанно углубляясь, идет от анимального полюса по окружности яйцеклетки по направлению к противолежащему вегетативному полюсу. Подобные борозды, проходящие по обоим полюсам шаровидной яйцеклетки наподобие меридианов на земном шаре, называются меридиональными бороздами.
Первая меридиональная борозда настолько глубоко проникает вглубь яйца, что из первоначально одной клетки возникают первые два бластомера, которые остаются вместе, прочно склеенными между собой. По истечении определенного времени наступает второе дробящее деление, при котором на поверхности обоих первых бластомеров возникает вторая, опять-таки меридиональная, борозда, идущая от анимального полюса к полюсу вегетативному, но перпендикулярно к плоскости хода первой борозды. Таким образом, из оплодотворенной яйцеклетки возникают четыре бластомера.
После кратковременной стадии покоя следует третье дробящее деление, которое проходит в плоскости, перпендикулярной к плоскостям обеих предыдущих борозд. Возникшая при этом борозда охватывает все четыре бластомера приблизительно на уровне экваториальной плоскости, лежащей между обоими полюсами. В соответствии со своим ходом данная борозда носит название широтной, или экваториальной, борозды. Благодаря третьему дроблению количество бластомеров вновь возрастает вдвое, так что маленький зародыш ланцетника на этой стадии развития имеет уже восемь клеток (бластомеров). Верхние четыре бластомера, лежащие над экваториальной бороздой в области анимального полюса, по размерам несколько меньше бластомеров, лежащих в области вегетативного полюса.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Бластогенез ланцетника. Дробление яйцеклетки ланцетника
Содержание темы
2. а) В разделе 3 (темы 5-6) мы уже рассматривали со сравнительных позиций ранние стадии развития различных животных - от ланцетника до млекопитающих.
б) Это дало нам общие представления о происхождении различных тканей и органов.
3. а) Теперь же (в теме 31) дадим последовательное описание ранних этапов развития человека - с привязкой этих этапов к конкретным срокам беременности и конкретному "месту действия".
б) Затем (в теме 32) будут рассмотрены строение и функционирование внезародышевых органов - в первую очередь, плаценты.
31.1. Периодика внутриутробного развития человека
2. а) Как видно, в эмбриогенезе человека различают 3 периода:
б) Заметим, что существуют и иные подразделения эмбриогенеза на периоды:
| зародышевый - первые 3 недели, | начальный - 1-я неделя, |
| эмбриональный - 4-8 недели, | зародышевый - 2-8 недели, |
| плодный - 9-40 недели. | плодный - 9-40 недели. |
в) Таким образом, разночтения касаются первых восьми недель развития.
3. а) Их-то мы и будем рассматривать в этой теме.
31.2. Половые клетки
31.2.1.1. Особенности строения
имеет объём 2-3 мл и
содержит 200-350 млн сперматозоидов ( Сз).
2. а) Подробно строение Сз излагалось в п. 5.1.2.
б) Напомним: в Сз различают 2 части -
в) При этом у человека головка Сз сильно уплощена.
таксисе Сз и
связывании с яйцеклеткой.
2. а) Заметим: эти белки (по крайней мере, те, что участвуют в связывании с яйцеклеткой) имеют
б) Узкой ножкой с мешочком связан также небольшой акросомный пузырёк (2а).
р езко уплотнено и
содержит гаплоидный набор хромосом.
б) О т одной из них начинается аксонема (7), или осевая нить хвоста, образованная микротрубочками по схеме
9 наружных фибрилл,
митохондриальная спиральная оболочка (6) и
фибриллярное влагалище (8) (9 наружных фибрилл и волокнистая оболочка), а также
31.2.1.2. Детерминация пола ребёнка
у 50 % Сз имеется Х-хромосома ,
а у других 50 % Сз - Y-хромосома .
2. Пол ребёнка определяется "полом" сперматозоида:
31.2.1.3. Препарат
На снимке (который нам знаком по п. 5.1.2.1) мы видим основные компоненты сперматозоида:
головку (1) и хвост (2),
31.2.2.1. Условность термина "яйцеклетка"
ооциты I и затем
(непосредственно перед овуляцией) ооциты II.
окружён блестящей и зернистой оболочками и
находится на стадии метафазы второго деления созревания.
происходят последние стадии этого деления (метафаза, анафаза и телофаза);
в результате, ядро ооцита II делится на 2 ядра с гаплоидными наборами хромосом,
и одно из этих ядер остаётся в качестве женского пронуклеуса (сосуществующего с мужским пронуклеусом),
так что из ооцита II, минуя стадию собственно яйцеклетки, образуется сразу зигота.
ооцит I (после стадии роста),
ооцит II или
только что образовавшуюся зиготу.
31.2.2.2. Специфические структуры цитоплазмы
I. Содержание желтка
в цитоплазме (1) равномерно распределено относительно небольшое количество желтка;
причём, в эволюции это вторично : впервые такой тип яйцеклетки встречается у ланцентника.
объём яйцеклетки человека в несколько тысяч раз больше объёма сперматозоида.
II. Перечень специфических структур
Специфические структуры цитоплазмы яйцеклетки (известные нам по п. 5.1.3.6) суммированы в таблице. -
после оплодотворения участвуют в кортикальной реакции (см. ниже - п. 31.3.1.2.III).
31.2.2.3. Другие особенности строения
Яйцеклетке присущи также следующие особенности. –
б) Отсутств уют центриол и;
в связи с этим, способность к делениям восстанавливается только тогда,
когда в клетку попадают центриоли сперматозоида.
блестящая, или прозрачная (zona pellucida, или Zp) ( 4 ) , и
зернистая ( 5 ) , образованная фолликулярны ми клетк ами.
а гликопротеины фракции Zp2 после кортикальной реакции
не окружены базальной мембраной (поскольку представляют собой лишь часть фолликулярного эпителия),
но имеют длинные отростки , пронизывающие блестящую оболочку.
либо только внутренние части фолликулярных клеток с отходящими от них отростками,
либо всю совокупность зернистого слоя и блестящей оболочки.
31.2.2.4. Препараты
Приведём два снимка из п. 5.1.3.4.
I. Окраска гематоксилин-эозином
б) Тем не менее, яйцеклетка имеет практически такое же строение.
2. На снимке видны следующие структуры :
ядро ооцита (1) и в нём - ядрышки (2),
цитоплазма ооцита с желточными гранулами (3),
блестящая (4) и зернистая (5) оболочк и,
соединительнотканная оболочка - тека ( 6 ).
II. Окраска по Маллори
2. Более отчётливо, чем при предыдущей окраске, видны следующие структуры:
ядро ооцита (1) и ядрышки (2) в нём,
цитоплазма ооцита (3),
блестящая (4) и зернистая (5) оболочки.
31.3. Зародышевый период развития
31.3.1.1. Общая характеристика
только один сперматозоид может проникнуть в яйцеклетку (точнее, ооцит II).
б) При этом в женских половых путях сперматозоиды сохраняют оплодотворяющую способность в течение 1-2 суток .
2. Оптимальный срок для оплодотворения - первые 24 часа после овуляции
(хотя ооцит II может сохранять способность к оплодотворению ещё некоторое время).
3. Таким образом, оплодотворение может наступить лишь в том случае, если половой акт совершается в интервал времени
момент овуляции + 1-2 суток
(минус - за счёт сохранения сперматозоидов в женских половых путях,
а плюс - за счёт сохранения ооцита II).
31.3.1.2. Основные события
Фазы оплодотворения были описаны в п. 5.2.2.
Кратко напомни м приводи вшуюся там информаци ю.
I. Сближение и дистантное взаимодействие половых клеток
это происходит пассивно - благодаря току слизи (выделяемой под действием эстрогенов).
б) Данный ток вызывается
биением ресничек мерцательных клеток и
тоническими сокращениями маточных труб (под действием прогестерона).
В матку они попадают, в основном, пассивно - благодаря тоническим сокращениям женских половых путей.
б) Затем часть Сз также, в основном, пассивно, достигает маточных труб.
2. Считается, что сокращения влагалища и матки усиливаются под влиянием простагландинов - гормональных компонентов спермы (синтезируемых в простате).
метаболизм и подвижность Сз резко усиливаются (п. 29.2.4.8).
а мембраны Сз в области головки теряют поверхностные гликопротеины и поэтому приобретают
способность связываться с блестящей оболочкой ооцита,
а также лабильн ость (что необходимо для последующего разрыва акросомы);
б) Вероятно, капацитацию инициируют
гиногамоны II , выделяемые ооцитом.
становится преимущественно активным
и обеспечивается биением их жгутиков .
б) При этом сперматозоиды одновременно
двигаются поступательно
и вращаются вокруг своей оси.
в) Направленность этого движения сперматозоидов обеспечивается
реотаксисом - способностью определять направление тока жидкости (в данном случае, слизи маточных труб) и двигаться против него),
а также хемотаксисом (способностью определять градиент концентрации определённых веществ - аттрактантов - и двигаться против него).
г) Считают, что такими аттрактантами являются
определённые пептиды, выделяемые ооцитом II или его окружением.
II. Контактное взаимодействие половых клеток
а) разрываются передние участки плазмолеммы и мембраны акросомы,
б) отчего высвобождаются акросомальные ферменты :
гиалуронидаза разъединяет клетки зернистой оболочки ,
а трипсиноподобный фермент акрозин и ряд других ферментов растворяют блестящую оболочку в месте прохождения Сз .
III. Проникновение сперматозоида в ооцит II
часть плазмолеммы Сз встраивается в мембрану ооцита,
а в ооцит проникают ядро ( 2 ) Сз и центриоли.
благодаря ионным каналам встроенной мембраны Сз, изменяется трансмембранный потенциал ооцита,
что стимулирует выброс содержимого кортикальных гранул (3) за пределы клетки.
б) Под влиянием выделяемых веществ
мембрана ооцита теряет рецепторную активность (модифицируются рецепторные гликопротеины Zp3 );
создаётся перивителлярное пространство (4) - между плазмолеммой и блестящей оболочкой (т.к. сюда привлекается вода ),
блестящая оболочка уплотняется (за счёт перестройки гликопротеинов Zp2 ) - образуется оболочка оплодотворения .
в) Всё это препятствует проникновению в ооцит II других Сз.
г) Кроме того, ооцитом выделяются
гиногамоны I, которые вызывают агглютинацию оставшихся сперматозоидов.
IV. Подготовка зиготы к дроблению
набухает (превращаясь в мужской пронуклеус (2) )
и сближается с женским пронуклеусом (сближенные ядра называются синкарионом ),
молекулы ДНК (в пронуклеусах) и
пришедшие с Сз центриоли (6) .
V. Начало первого митотического деления
а) их оболочки разрушаются,
б) а хромосомы
конденсируются и
в метафазе образуют единую материнскую звезду (7).
Заметим: всё это время продолжается медленное пассивное продвижение ооцита II, а затем зиготы, по яйцеводу к матке.
31.3.1.3. Дополнительная иллюстрация
она не имеет клеточной структуры, т.к. происходит из блестящей оболочки .
2. а) Внутри зиготы - два ядра-пронуклеуса (2) почти равного объёма.
б) Поскольку ядра уже соприкасаются (что является сигналом к началу митоза), в них
уже совершилось удвоение ДНК ,
и хромосомы стали двухроматидными.
31.3.2.1. Общая характеристика
б) Рост клеток затруднён оттого, что вокруг зародыша сохраняется плотная оболочка оплодотворения (1), которая препятствует
и притоку питательных веществ извне,
(жизнедеятельность поддерживается за счёт расходования резервов яйцеклетки);и самому увеличению размера зародыша.
образуются всё более мелкие клетки и
общий объём зародыша не увеличивается .
Дробление происходит в просвете яйцевода,
и к концу его зародыш достигает (продвигаясь по яйцеводу) полости матки.
полное: дробятся все клетки зародыша ;
асинхронное : клетки делятся не одновременно; поэтому могут быть стадии с нечётным количеством бластомеров;
неравномерное: образуются клетки разного размера.
б) Поэтому при диссоциации клеток на этих стадиях каждый бластомер даёт начало самостоятельному зародышу,
чем и объясняется появление однояйцевых близнецов.
всё более различаются друг от друга по виду и потенциям развития,
а вместе с тем теряют и свойство тотипотентности (способность развиваться в отдельный организм).
В т.ч. в центре находятся 3-4 тёмные и крупные клетки - предшественники эмбриобласта.
Остальные, периферические, клетки - светлые и мелкие; это предшественники трофобласта.
31.3.2.2. Динамика увеличения числа клеток
первое деление дробления завершается через 30 часов,
а всего на стадии дробления происходит 5 циклов делений (2 5 = 32).
31.3.3. Образование бластоцисты
31.3.3.1. Общая характеристика
зародышевый пузырёк, заполненный жидкостью.
2. В вид е свободной бластоцисты зародыш находится в полости матки около 2-х суток -
3. Деления клеток в бластоцисте по-прежнему являются
б) В самих бластомерах всё более активируются синтетические процессы.
2 . а) В трофобласте появляются выросты,
которые постепенно разрушают оболочку оплодотворения вокруг зародыша.
б) В результате, за несколько часов до имплантации (см. ниже) зародыш теряет эту оболочку.
3. а) После этого оболочка уже не мешает зародышу увеличиваться в размере,
и с этих пор митотические циклы клеток становятся обычными, т.е. включают фазу роста.
Эмбриональный период на примере ланцетника. Бластогенез ланцетника
Онтогенезом, или индивидуальным развитием, называют весь период жизни особи с момента слияния сперматозоидов с яйцом и образования зиготы до гибели организма. Онтогенез делится на два периода: 1) эмбриональный – от образования зиготы до рождения или же выхода из яйцевых оболочек; 2) постэмбриональный – от выхода из яйцевых оболочек или рождения до смерти организма.
У большинства многоклеточных животных стадии эмбрионального развития, которые проходит зародыш, едины. В эмбриональном периоде выделяют три основных этапа: дробление, гаструляцию и первичный органогенез.
Развитие организма начинается с одноклеточной стадии. В результате многократных делений одноклеточный организм превращается в многоклеточный. Образующиеся клетки называются бластомерами. При делении бластомеров размеры их не увеличиваются, поэтому процесс деления носит название дробления. В период дробления накапливается клеточный материал для дальнейшего развития.
По мере увеличения числа клеток деление их становится неодновременным. Бластомеры все дальше и дальше отходят от центра зародыша, образуя полость – бластоцель. Завершается дробление образование однослойного многоклеточного зародыша – бластулы.
Особенностью дробления является чрезвычайно короткий митотический цикл бластомеров по сравнению с клетками взрослого организма. Во время очень короткой интерфазы происходит только удвоение ДНК.
Бластула, как правило, состоящая из большого числа бластомеров (у ланцетника – из 3000 клеток), в процессе развития переходит в новую стадию, которая называется гаструлой. Зародыш на этой стадии состоит из разделенных пластов клеток, так называемых зародышевых листков: наружного, или эктодермы, и внутреннего, или энтодермы. Совокупность процессов, приводящих к образованию гаструлы, называется гаструляцией. У ланцетника гаструляция осуществляется путем впячивания части стенки бластулы в первичную полость тела.
После завершения гаструляции у зародыша образуется комплекс осевых органов: нервная трубка, хорда, кишечная трубка. Эктодерма прогибается, превращаясь в желобок, а энтодерма, расположенная справа и слева от него, начинает нарастать на его края. Желобок погружается под энтодерму, и края его смыкаются. Образуется нервная трубка. Вся остальная эктодерма – зачаток кожного эпителия. На этой стадии зародыш носит название нейрулы.
Спинная часть энтодермы, располагающаяся непосредственно под нервным зачатком, обособляется от остальной энтодермы и сворачивается в плотный тяж – хорду. Из оставшейся части энтодермы развивается мезодерма и эпителий кишечника. Дальнейшая дифференцировка клеток зародыша приводит к возникновению многочисленных производных зародышевых листков – органов и тканей.
Из эктодермы развивается нервная система, эпидермис кожи и его производные, эпителий, выстилающий внутренние органы. Из энтодермы развиваются эпителиальные ткани, выстилающие пищевод, желудок, кишечник, дыхательные пути, печень, поджелудочную железу, эпителий желчного и мочевого пузыря, мочеиспускательного канала, щитовидную и околощитовидную железы.
Производными мезодермы являются: дерма, вся собственно соединительная ткань, кости скелета, хрящи, кровеносная и лимфатическая системы, дентин зубов, почки, половые железы, мускулатура.
Зародыш животных развивается как единый организм, в котором все клетки, ткани и органы находятся в тесном взаимодействии. При этом один зачаток оказывает влияние на другой, в значительной мере определяя путь его развития. Кроме того, на темпы роста и развития зародыша оказывают влияние внешние и внутренние условия.
Позвоночные животные произошли от бесчерепных. Современным представителем подтипа бесчерепных является ланцетник (рис. 34). В развитии ланцетника мы видим наиболее простую схему зародышевого развития хордовых животных, которая в значительной степени усложнилась в процессе эволюции у позвоночных животных и особенно у человека.
Ланцетник - морское животное. Самка и самец выбрасывают половые клетки (яйца и сперматозоиды) непосредственно в воду, где и происходит оплодотворение и развитие зародыша.
Вслед за оплодотворением зигота вступает в период дробления (рис. 35); количество бластомеров быстро увеличивается (2, 4, 8, 16 и т. д.).

В процессе деления бластомеры постепенно отодвигаются от центра зародыша к периферии, образуя в центре все увеличивающуюся полость.
В связи с этим к концу периода дробления возникает типичная бластула, стенка которой образована одним слоем клеток (бластодерма), а полость ее (бластоцель) заполнена жидкостью. Следующий период (гаструляция) связан с инвагинацией, т. е. впячиванием одной (вегетативной) половины бластулы в другую (анимальную) * (рис. 35). В результате образуется гаструла ** , у которой имеется внутренний зародышевый листок (первичная энтодерма), гастроцель (полость первичной кишки) и бластопор (первичный рот).
* (В яйце ланцетника в одной из его половин содержится больше желтка, чем в другой. В результате изучения развития зародышей установлено, что часть яйца, снабженная большим количеством желтка, при дроблении образует ту половину бластодермы, которая впячивается в периоде гаструляции и образует внутренний зародышевый листок - энтодерму. Известно, что из энтодермы образуется пищеварительная и другие системы так называемой растительной (вегетативной) жизни. Поэтому и часть яйца, содержащая большее количество желтка, и образующаяся из него в результате дробления часть бластулы называются вегетативными частями. Из противоположной части яйца и соответствующей части бластулы развиваются эктодерма и далее органы анимальной (животной) части, в частности нервная система и др. Поэтому эти части яйца и бластулы именуются анимальными. )
** (Греч, gaster - желудок. Отсюда название "гаструла", чтобы подчеркнуть, что зародыш в этой стадии снабжен зачатком пищеварительной системы в виде первичной кишки. )
Бластоцель (первичная полость тела) в период инвагинации сохраняется некоторое время в виде узкой щели между наружным и внутренним зародышевыми листками, а затем исчезает. После сформирования гаструла начинает увеличиваться в длину; одновременно происходит концентрическое смыкание краев бластопора (первичного рта) * .
* (Бластопор (первичный рот), соединяющий первичную кишку с внешней средой, у одних животных на последующих этапах развития остается в качестве ротового отверстия (первичноротые животные), а у других становится заднепроходным (анальным) отверстием (вторичноротые животные). В последнем случае ротовое отверстие образуется на противоположном конце. К первичноротым относятся черви, моллюски и членистоногие, к вторичноротым - иглокожие и хордовые, в частности ланцетник и позвоночные животные, включая человека. )
Окончание гаструляции совпадает с началом периода обособления основных зачатков органов и тканей (рис. 36). В это время утолщенный дорсальный участок первичной эктодермы превращается в нервную пластинку, из которой возникает, проходя через стадию нервного желобка, нервная трубка (рис. 36, 37). Нервная трубка является зачатком нервной системы.


Другая часть наружного зародышевого листка при дальнейшем развитии замыкается над нервной трубкой и является зачатком кожного эпителия (эпидермиса).
Внутренний зародышевый листок претерпевает ряд изменений в клеточном составе, затем из него обособляются следующие образования: в области средней части его крыши - хордальная пластинка, из которой образуется зачаток хорды; в области латеральных частей крыши первичной кишки - карманообразные выпячивания, которые затем обособляются от стенки первичной кишки. Клеточный материал отшнуровавшихся карманообразных выпячиваний первичной кишки заполняет первичную полость тела (находящуюся между эктодермой и энтодермой) и представляет собой зачаток среднего зародышевого листка (мезодермы). В центре обособившегося участка оказывается отшнуровавшееся от полости первичной кишки пространство, являющееся вторичной полостью тела. Оставшаяся часть первичной энтодермы (дно первичной кишки) образует кишечную трубку, которая является зачатком вторичной (окончательной) кишки и из которой в дальнейшем возникает кишечный эпителий.
Таким образом, к концу гаструляции, в период обособления зачатков органов и тканей, нервная трубка оказывается расположенной на спинной стороне зародыша в срединном положении, а под ней последовательно расположены хорда и кишечная трубка. Окончательно выявляется билатеральная симметрия. Появляются мезодермальные карманы. Зачатки мезодермы с полостями внутри них врастают справа и слева в щель между кожной эктодермой и кишечной трубкой и соединяются под последней. В то же время мезодерма, вытянутая вдоль тела между кожной эктодермой, с одной стороны, хордой и кишечной трубкой, с другой стороны, разделяется на ряд отдельных участков (сегментов), расположенных по длине тела друг около друга. Сегментации подвергаются левый и правый мезодермальные карманы. Одновременно каждый мезодермальный мешок на всем протяжении делится на дорсальный участок (сомит) и вентральный участок (спланхнотом). Сомиты утрачивают полость, становятся плотными и служат исходным материалом для туловищной мускулатуры. Спланхнотомы сохраняют полость. Они остаются некоторое время отделенными друг от друга (в результате сегментации), а затем изолированные полости, заключенные в отдельных спланхнотомах, сливаются, так что образуется единая для всех сегментов тела целомическая полость (вторичная полость тела, целом). Материал стенок спланхнотомов является исходным материалом эпителиальной выстилки вторичной полости тела * .
* (Вторичная полость тела (целом) возникает в толще среднего зародышевого листка. Из нее в процессе эмбрионального развития образуются различные полости. У человека таковыми являются, в частности, брюшинная и плевральные полости и полость перикарда. Первичная полость тела (бластоцель) исчезает в процессе гаструляции и образования мезодермы. )
Развитие личинки ланцетника заканчивается появлением ротового и анального отверстий, жаберных щелей, формированием органов и др.
Из боковой пластинки развиваются брюшина, брыжейки (в которых в виде продольных каналов возникают основные кровеносные сосуды), мускулатура кишечника. Нефридиальные канальцы развиваются в виде пальцевидных выпячиваний стенок вторичной полости тела. Гонады развиваются как выпячивания той части стенок полости тела, которая соответствует месту разделения сомита и боковой пластинки гонотому. Рот образуется путем выпячивания первичной кишки на конце, противоположном гастропору (первичному рту), и встречного впячивания эктодермы. В месте встречи этих образований возникает прорыв. Закладка рта и жаберных щелей происходит асимметрично. Ротовое отверстие закладывается на нижней левой стороне зародыша. Левые жаберные щели (их 14) первоначально возникают на брюшной стороне, а затем перемещаются на правую сторону зародыша. Затем здесь появляется еще один ряд щелей (их 8), расположенных выше упомянутых ранее 14 щелей. Впоследствии нижний ряд щелей смещается на брюшную сторону и лишь после этого - на левую сторону тела. Число их при этом сокращается с 14 до 8. Число жаберных щелей с обеих сторон затем резко возрастает. Впоследствии рот смещается на брюшную сторону. Атриальная полость возникает первоначально в виде желобка на нижней поверхности тела. Формирующие этот желобок метаплевральные складки растут навстречу друг другу и, смыкаясь, образуют полость, открывающуюся наружу лишь в задней своей части, где упомянутые складки не срастаются. В целом личиночное развитие ланцетника длится около трех месяцев.
Дробление приводит к образованию шарообразного зародыша – бластулы . Если образуется сплошной шар без полости внутри, то такой зародыш называют морулой. Образование бластулы или морулы зависит от свойств цитоплазмы. Бластула образуется при достаточной вязкости цитоплазмы, морула – при слабой вязкости. При достаточной вязкости цитоплазмы бластомеры сохраняют округлую форму и только в местах соприкосновения слегка сплющиваются. Вследствие этого между ними появляется щель, которая по мере дробления увеличивается, заполняется жидкостью и превращается в бластоцель. При слабой вязкости цитоплазмы бластомеры не округляются и располагаются тесно друг возле друга, щели нет и полость не образуется. Бластулы различны по своему строению и зависят от .
Типы бластул
Различают пять типов бластул : целобластулу, амфибластулу, стерробластулу, дискобластулу и перибластулу. Целобластула образуется при полном равномерном из яйцеклеток гомолецитального типа (ланцетник). Бластодерма целобластулы состоит из одного ряда более или менее одинаковых бластомеров, внутри находится крупная полость – бластоцель.
Бластодерма амфибластулы состоит из нескольких рядов клеток. Бластодерма в анимальной части тоньше, чем в вегетативной. Бластоцель меньших размеров, чем у , и смещена к анимальному полюсу. Такого типа бластула образуется при полном неравномерном дроблении и характерна для круглоротых и .
Стерробластула состоит из одного ряда крупных бластомеров, которые глубоко заходят в полость бластулы, бластоцель в связи с этим или очень малая, или отсутствует (некоторые членистоногие).
Дискобластула образуется при неполном дискоидальном дроблении. Бластоцель в виде узкой щели находится между зародышевым диском и желтком. Крыша бластулы представлена бластодермой, а дно желтком. Такая бластула характерна для костистых , пресмыкающихся и птиц. Бластодерма перибластулы состоит из одного ряда клеток, которые окружают желток. Полость в ней отсутствует. Перибластула наблюдается у некоторых насекомых.
Результат активного деления клеток, роста и направленных перемещений (миграций) клеточных потоков с формированием многослойного зародыша, или гаструлы, (возникновением послойно расположенных, отделенных друг от друга отчетливой щелью, зародышевых листков: наружного - эктодермы, среднего - мезодермы, внутреннего - энтодермы). Перемещение клеток происходит в строго определенной области зародыша - в области серого серпа. Последний был описан В. Ру в 1888 г. В оплодотворенном яйце амфибии серый серп выявляется как окрашенная область на стороне, противоположной проникновению спермия. В этом месте, как полагают, локализуются факторы, необходимые для гаструляции.
У разных представителей позвоночных совершается несколькими основными способами: путем инвагинации (впячивания), иммиграции (перемещения части клеток внутрь зародыша), эпиболии (обрастания), деламинации (расщепления). Способы гаструляции зависят от типа яйцеклетки. При любом способе гаструляции в качестве ведущих сил выступают неравномерная пролиферация клеток в разных частях зародыша, уровень обменных процессов в клетках, расположенных в разных частях зародыша, активность амебоидных движений клеток, а также индуктивные факторы (белки, нуклеопротеиды, стероиды и др.). В результате гаструляции обособляются основные зачатки органов и тканей.
Следующим периодом эмбриогенеза является гисто- и органогенез - дифференцировка различных тканей и органов организма из материала зародышевых листков и эмбриональных зачатков.
В результате гаструляции возникает многослойный зародыш . Несмотря на различные способы гаструляции после выделения материала зародышевых листков по оси зародыша находится материал хорды, который подстилает нервную пластинку, слева и справа от хорды располагается материал мезодермы. Все это характеризует осевой комплекс зачатков. В дальнейшем происходит формирование зачатков органов, представляющих собой пространственно локализованные группы стволовых клеток - источников развития тканей. Закономерности дифференцировки клеточного материала зачатков можно проследить в эмбриогенезе наиболее изученных животных.
Ланцетник. Развитие ланцетника.
Классическим объектом эмбриологических исследований ланцентник , подробно изученный А.О. Ковалевским. Ланцетник является представителем класса хордовых животных подтипа бесчерепных, размером до 8 см и обитает на песчаном дне в теплых морях. Свое название получил из-за формы, напоминающей ланцет (хирургический инструмент с обоюдоострым лезвием, современный скальпель).
Яйцеклетка ланцетника олиго- и изолецитальная, размером 110 мкм, ядро располагается ближе к анимальному полюсу. Оплодотворение наружное. Дробление зиготы полное, почти равномерное, синхронное и заканчивается образованием бластулы. В результате чередования меридианальных и широтных борозд дробления формируется однослойная бластула с полостью, заполненной жидкостью - бластоцелем. Бластула сохраняет полярность, ее дно представляет собой вегетативную, а крыша - анимальную части; между ними находится краевая зона.
При гаструляции происходит впячивание вегетативной части бластулы в анимальную. Впячивание постепенно углубляется и, наконец, формируется двустенная чаша с широко зияющим отверстием, ведущим в новообразованную полость зародыша. Такой способ гаструляции называется инвагинацией. Так бластула превращается в гаструлу. В ней материал зародыша оказывается дифференцированным на наружный листок - эктодерму, и внутренний - энтодерму. Полость чаши именуется гастроцелем, или полостью первичной кишки, которая сообщается с внешней средой посредством бластопора, что соответствует заднепроходному отверстию. В бластопоре различают дорсальную, вентральную и две латеральных губы. В результате инвагинации центр тяжести зародыша смещается, и зародыш поворачивается бластопором кверху. Постепенно происходит смыкание краев бластопора и удлинение зародыша. Топография клеток в составе губ бластопора определяет развитие разных частей зародыша. При гаструляции из внутреннего листка гаструлы обособляются хорда и мезодерма, которые располагаются между экто- и энтодермой. Завершается гаструляция образованием осевого комплекса зачатков и дальше - обособлением зачатков органов. Хорда индуцирует развитие нервной трубки из материала дорсальной эктодермы. Эта часть эктодермы утолщается, образуется нервная пластинка (нейроэктодерма), которая прогибается по средней линии и превращается в желобок.

Методы маркирования для изучения процессов миграции бластомеров позволили выделить определенные области зародыша на ранних стадиях развития (зиготы - бластулы), которые позднее развиваются в зародышевые листки и эмбриональные зачатки органов и тканей. Эти области были названы презумптивными (предполагаемыми) участками, или зачатками.
Эмбриогенез ланцетника
Индивидуальное развитие ланцетника представляет собой простейшую исходную схему эмбриогенеза, путем постепенного усложнения которой в ходе эволюции возникли более сложные системы развития хордовых животных, включая человека.
СТРОЕНИЕ ЯЙЦЕКЛЕТКИ. ОПЛОДОТВОРЕНИЕ
Яйца ланцетника бедны желтком и микроскопически малы (100-120мкм), относятся к изолецитальному типу. Желточные зерна невелики и распределены в цитоплазме почти равномерно. Тем не менее, в яйцеклетке выделяют анимальный и вегетативный полюс. В области анимального полюса при созревании яйцеклетки происходит отделение редукционных телец. Ядро в оплодотворенной яйцеклетке находится ближе к анимальному полюсу в силу не совсем равномерного распределения желтка, располагаясь в свободной от желточных включений части клетки. Созревание яйцеклетки происходит в воде. Первое редукционное тельце отделяется на анимальном полюсе ооцита еще до оплодотворения. Оно смывается водой и погибает.
Самки ланцетника выметывают яйца в воду, сюда же самцы выпускают сперматозоиды – оплодотворение наружное, моноспермное. После проникновения сперматозоида вокруг яйцеклетки образуется оболочка оплодотворения, которая препятствует проникновению в яйцо других
избыточных спермиев. Вслед за этим происходит отделение второго редукционного тельца, которое располагается между желточной оболочкой и яйцеклеткой.
Все дальнейшее развитие проходит также в воде. Через 4-5 дней из яйцевой оболочки вылупливается микроскопическая личинка, которая переходит к самостоятельному питанию. Сначала она плавает, а затем оседает на дно, растет и совершает метаморфоз
ДРОБЛЕНИЕ. БЛАСТУЛА
Малое количество желтка объясняет простоту дробления и гаструляции. Дробление полное, почти равномерное, радиального типа, в результате образуется целобластула (рис. 1).
Рис. 1. Дробление яйца ланцетника (по Алмазову, Сутулову, 1978):
А – зигота; Б, В, Г – образование бластомеров (показано расположение веретена деления)
Анимальный полюс приблизительно соответствует будущему переднему концу тела личинки. Оплодотворенная яйцеклетка (зигота) целиком дробится на бластомеры в правильной геометрической прогрессии. Бластомеры почти одинаковой величины, анимальные лишь не-
сколько мельче вегетативных. Первая борозда дробления – меридиональная, проходит через анимальный и вегетативный полюс. Она разделяет шаровидное яйцо на две совершенно симметричные половины, но бластомеры округляются. Они шаровидные, имеют малую площадь со-
прикосновения. Вторая борозда дробления также меридиональная, перпендикулярна первой, а третья – широтная.
По мере увеличения количества бластомеров они все больше расходятся от центра зародыша, образуя посередине большую полость. В конце концов, зародыш принимает форму типичной целобластулы – пузырька со стенкой, образованной одним слоем клеток – бластодермой и с полостью, заполненной жидкостью – бластоцелем (рис. 2).
Клетки бластулы, вначале округлые и потому не плотно сомкнутые, затем приобретают форму призм и плотно смыкаются. Поэтому позднюю бластулу, в противоположность ранней, называют эпителиальной.
Стадия поздней бластулы завершает период дробления. К концу этого периода размеры клеток достигают минимума, а общая масса зародыша не увеличивается по сравнению с массой оплодотворенной яйцеклетки.


Рис. 2. Бластула ланцетника (по Алмазову, Сутулову, 1978):
А – внешний вид; Б – поперечный разрез (стрелка показывает задне-переднее направление тела будущего зародыша); В – расположение материалов будущих органов на сагитальном разрезе бластулы
Гаструляция происходит путем инвагинации – впячивания вегетативного полушария бластулы внутрь, по направлению к анимальному полюсу (рис. 3). Процесс протекает постепенно и заканчивается тем, что все вегетативное полушарие бластулы уходит внутрь и становится внутренним зародышевым листком – первичная энтодерма зародыша. Фактор, обуславливающий инвагинацию – разница темпов деления клеток в краевой зоне и в вегетативной части бластулы, приводящей к активному перемещению клеточного материала. Анимальное полушарие становится

Рис. 3. Начальные стадии гаструляции ланцетника (по Мануиловой, 1973):
наружным зародышевым листком – первичная эктодерма. Зародыш приобретает вид двуслойной чаши с широко зияющим отверстием – первичным ртом или бластопором. Полость, в которую ведет бластопор, называют гастроцель (полость первичного кишечника). Бластоцель в результате впячивания низводится до узкой щели между наружным и внутренним зародышевыми листками. На данной стадии зародыш носит название гаструлы (рис. 4 А, Б).
Первичный кишечник (архентерон), представленный внутренним зародышевым листком, окружающим полость гаструлы, является зачатком не только пищеварительной системы, но и других органов и тканей личинки. Бластула, как и яйцеклетка, плавает анимальным полюсом кверху в силу большего веса вегетативного полушария.
В результате инвагинации центр тяжести зародыша перемещается и гаструла поворачивается бластопором кверху.
Бластопор окружен дорсальной, вентральной и боковыми губами. Далее происходит концентрическое смыкание краев бластопора и удлинение зародыша. У ланцетника, представителя вторичноротых, бластопор соответствует не ротовому, а заднепроходному отверстию, обозначая
задний конец зародыша. В результате смыкания краев бластопора и выпячивания тела в передне-заднем направлении, зародыш удлиняется. При этом поперечник гаструлы уменьшается – общая масса составляющих зародыш клеток не может увеличиваться, пока развитие идет под покровом яйцевых оболочек. Зародыш приобретает билатеральную симметрию.
Расположение зачатков в поздней гаструле лучше всего видно на поперечном разрезе зародыша (рис. 4 В, Г).
Наружную стенку его образует эктодерма, неоднородная в своем составе. В дорcальной части эктодерма утолщена и состоит из высоких цилиндрических клеток. Это зачаток нервной системы, которая остается


Гаструла ланцетника (по Мануиловой, 1973):
А – ранняя стадия; Б – поздняя стадия; В – поперечный разрез через позднюю гаструлу; Г – гаструла, переходящая в нейрулу (поперечный разрез)
еще на поверхности и образует так называемую медуллярную или нервную пластинку. Остальная эктодерма состоит из мелких клеток и является зачатком покрова животного. Под нервной пластинкой во внутреннем зародышевом листке располагается зачаток хорды, по обеим сторонам которого в виде двух тяжей находится материал мезодермы. В брюшной части располагается энтодерма, образующая основание первичной кишки, крышу которой составляют зачатки хорды и мезодермы .
Материал будущих внутренних органов, находясь в бластуле снаружи, в процессе гаструляции перемещается внутрь зародыша и располагается на местах, развивающихся из них органов. Только зачаток нервной системы остается еще на поверхности. Он погружается внутрь зародыша на стадии, следующей за гаструлой.
НЕЙРУЛЯЦИЯ И ОБРАЗОВАНИЕ ОСЕВЫХ ОРГАНОВ
По окончании гаструляции начинается следующий этап в развитии зародыша – дифференцировка зародышевых листков и закладка органов. Наличие комплекса спинных органов: нервной трубки, хорды и осевой мускулатуры, известных также под именем осевых, является одной их
характерных черт типа хордовых.
Стадия, на которой происходит закладка осевых органов, называется нейрулой. Внешне она характеризуется изменениями, происходящими с зачатком нервной системы.
Они начинаются с нарастания эктодермы по краям нервной пластинки. Образующиеся нервные валики растут навстречу друг другу и затем смыкаются. Пластинка же погружается внутрь и сильно прогибается (рис. 5).

Рис. 5. Нейрула ланцетника (по Мануиловой, 1973):
А – ранняя стадия (поперечный разрез); Б – поздняя стадия (поперечный
разрез), буквой “ Ц ” обозначена вторичная полость тела (целом)
Это приводит к образованию желобка, а затем нервной трубки, которая в передней и задней части зародыша некоторое время остается открытой (указанные изменения удобнее всего проследить на поперечном разрезе зародыша). Вскоре, в задней части тела эктодерма нарастает на бластопор и отверстие нервной трубки, закрывая их таким образом, что нервная трубка остается сообщенной с кишечной полостью – образуется нервно-кишечный канал.
Одновременно с формированием нервной трубки существенные изменения происходят и во внутреннем зародышевом листке. Из него постепенно обособляются материалы будущих внутренних органов. Зачаток хорды начинает выгибаться, выделяется из общей пластинки и превращается в обособленный тяж в виде сплошного цилиндра. Одновременно происходит обособление мезодермы. Этот процесс начинается с появления небольших карманообразных выростов по двум сторонам
внутреннего листка. По мере роста они отделяются от энтодермы и в ви-де двух тяжей с полостью внутри располагаются по всей длине зародыша. Кроме продольных желобков от переднего конца первичного кишечника последовательно отчленяются еще две пары целомических мешков.
Таким образом, в развитии ланцетника имеется стадия, характеризующаяся наличием трех пар сегментов и свидетельствующая об эволюционном родстве ланцетника с трехсегментарными личинками полухордовых и иглокожих. У ланцетника ярко выражен энтероцельный способ
образования целома – его отшнуровка от первичного кишечника. Этот способ является исходным для всех вторичноротых животных, но почти ни у кого из вышестоящих позвоночных, за исключением круглоротых, с такой ясностью не представлен. После отделения хорды и мезодермы
края энтодермы постепенно сближаются в спинной части и в конце концов смыкаются, образуя замкнутую кишечную трубку.
В ходе дальнейшего развития мезодерма сегментируется: тяжи разделяются поперечно на первичные сегменты или сомиты. Из них образуются три основные закладки:
• дерматом формируется из наружной, обращенной к эктодерме стенке сомита, – из его клеток впоследствие возникает соединительная часть кожи, представленная преимущественно фибробластами;
• склеротом образуется из внутренней части сомита, примыкающей к хорде (низшие позвоночные) или к хорде и нервной трубке (высшие позвоночные) – представляет зачаток осевого скелета;
• миотом представляет часть сомита, расположенную между дерматомом и склеротомом – является зачатком всей поперечно-полосатой мускулатуры.
Дифференцировка сомитов у ланцетника протекает иначе, чем у позвоночных. Это различие выражается в том, что у позвоночных сегментируется только спинная часть мезодермальных тяжей, тогда как у ланцетника они полностью распадаются на сегменты. Последние вскоре разделяются на спинную часть – сомиты, и брюшную – спланхнотом.
Сомиты, из которых развивается туловищная мускулатура, остаются обособленными друг от друга, спланхнотомы же сливаются на каждой стороне, образуя левую и правую полости, которые затем объединяются под кишечной трубкой в общую вторичную полость тела (целом).
В развитии ланцетника, с одной стороны, ясно представлены черты типичных позвоночных (характерное расположение зачатков при гаструляции, формирование хорды из дорсальной стенки первичной кишки и нервной пластинки из дорсальной эктодермы), а с другой – черты беспозвоночных вторичноротых животных (целобластула, инвагинационная гаструла, трехсегментная стадия, энтероцельная закладка мезодермы и образования целома).
В дальнейшем, в связи с образованием хвоста нервно-кишечный канал исчезает. В головной части кишечной трубки прорывается ротовое отверстие, а на заднем конце, под хвостом, образуется анальное – путем вторичного прорыва стенки тела животного на месте закрывшегося бластопора. Зародыш переходит в стадию свободноплавающей личинки.
Читайте также: