Эндоплазматический ретикулум и митохондрии. Клеточный цикл
Обновлено: 22.04.2024
Клеточная биология - раздел биологии, изучающий живые клетки, их органеллы, их строение, функционирование, процессы клеточного размножения, старения и смерти. Настоящая программа учебной дисциплины устанавливает требования к образовательным результатам и результатам обучения студента и определяет содержание и виды учебных занятий и отчетности. Программа предназначена для преподавателей, ведущих дисциплину «Клеточная биология: основы биопроцессов», учебных ассистентов и студентов направления подготовки 06.03.01 Биология, обучающихся по образовательной программе «Клеточная и молекулярная биотехнология». Программа учебной дисциплины разработана в соответствии с: Образовательным стандартом НИУ ВШЭ по направлению 06.03.01 Биология; Образовательной программой «Клеточная и молекулярная биотехнология» и объединенным учебным планом по образовательной программе «Клеточная и молекулярная биотехнология». Данная учебная дисциплина включена в раздел «Вариативная профессиональная часть» Учебного плана 06.03.01 Биология и относится к базовой профильной части. Осваивается на 1 курсе в 3-4 модуле. Изучение данной дисциплины базируется на результатах освоения дисциплины «Общая и неорганическая химия», «Зоология беспозвоночных». Основные положения дисциплины «Клеточная биология: основы биопроцессов» будут использоваться при прохождении дисциплин «Молекулярная биология», «Микробиология», «Биохимия», «Биофизика», «Генетика», «Клеточные технологии и инженерия», «Генная инженерия», «Основы молекулярной онкологии (онкоиммунология)» и практики на 1-4 курсах образовательной программы «Клеточная и молекулярная биотехнология».
Цель освоения дисциплины
Целью освоения дисциплины являются приобретение знаний в области клеточной биологии и химических основ биологических процессов, происходящих на уровне клетки, а также базовых навыков работы с клетками прокариотических и эукариотических организмов.
Планируемые результаты обучения
Сравнивает прокариоты и эукариоты на примере бактерий и дрожжей. Сопоставляет их с культивируемыми клетками человека.
Описывает основные элементы строения клеточных мембран. Описывает, как устроен трансмембранный транспорт в клетке и основные этапы везикулярного транспорта. Анализирует базовые механизмы локализации, биогенеза и секреции белков и решает типовые задачи на локализацию белков. Решает типовые задачи на клеточный транспорт.
Объясняет и иллюстрирует организацию хромосом в клетке, структуру хроматина и транскрипции. Формулирует, как устроено клеточное ядро, основные субъядерные структуры. Применяет эти знания для решения типовых задач.
Распознаёт органеллы клетки, обсуждает их функции и протекающие в них процессы. Описывает структуру и функции клеточных органелл, применяет эти знания для решения типовых задач клеточной биологии.
Описывает основные механизмы деления и гибели клеток, наблюдает в микроскоп деление и апоптоз эукариотической клетки. Описывает строение, состав и динамику цитоскелета. Решает типовые задачи, посвященные строению, делению и гибели клеток.
Описывает базовые механизмы внутриклеточной передачи сигнала и межклеточного взаимодействия. Знакомится с особенностями строения наиболее известных типов клеток человека.
Знакомится с базами данных NCBI и геномным UCSC. Создает простой запрос Blast и находит интересующий участок генома в геномном браузере. Создает запрос Pubchem для поиска низкомолекулярных соединений.
Описывает основные клеточные органеллы. Культивирует клеточные линиями млекопитающих и проводит прижизненное окрашивания и наблюдения клеток. Проводит функциональные клеточные тесты на примере скретч-теста и анализа цитотоксичности.
Содержание учебной дисциплины
Прокариотические клетки (бактерии и археи), их внутренняя организация. Грамм-положительные и отрицательные бактерии. Возникновение эукариот и их органелл. Эукариотические клетки и их внутренняя организация. Общий вид мембранных органелл: ядро, эндоплазматический ретикулум (ЭР), митохондрии и хлоропласты. Базовые элементы строения ДНК (структура нуклеотидов и двойная спираль), РНК (особенности её одноцепочечной структуры), белков (а-спираль и b-листы) .
Культивирование микроорганизмов. Жидкие и гелевые питательные среды. Селективные и маркерные среды. Оценка фенотипических эффектов. Клеточные культуры эукариот. Выделение клеток многоклеточных организмов в культуру. Клеточные культуры многоклеточных организмов: генотипирование и проблемы контаминации. Проточная цитометрия и сортировка клеток. Микроскопия: световая, флуоресцентная, флуоресцентная сверхвысокого разрешения, электронная. Понятие конфокальной микроскопии и трехмерной реконструкции. Приготовление образцов. Окрашивание низкомолекулярными флуорофорами и иммуноокрашивание. Понятие о флуоресцентных белках. Разрушение клеток, разделение клеточных компонентов центрифугированием.
Мембраны клеток. Мембранные органеллы эукариот. Особенности строения и состава мембран клеток и органелл. Липиды, фосфолипиды, холестерин. Липидные островки. Состав мембран разных клеток и органелл. Подвижность/текучесть мембраны и влияющие на нее факторы. Асимметрия сторон мембран. Гликолипиды. Мембранные белки. Ключевые структурные элементы. Способы локализация в мембране. Индекс гидрофобности участков белков. Сборка белковых комплексов в мембране на примере Т-клеточного рецептора. Регуляторная роль фосфоинозитидов. Ограничение подвижности белков в мембране. Понятие о цитоскелете кортикального слоя. Клеточные стенки бактерий, грибов, растений: краткий обзор строения.
Проницаемость мембраны для различных молекул. Мембранный транспорт: активный и пассивный. Ион-селективный пассивный мембранный транспорт Активный транспорт: АТФ-зависимый и сопряженный; понятие о светозависимом. Элементы строения белков-транспортеров. Понятие об АТФ-зависимых насосах. Эффлюкс антибиотиков бактериями через одиночную и двойную мембрану. Асимметрия расположения транспортёров в мембране на примере транспорта глюкозы. Мембранный транспорт: электрохимический градиент. Регуляция ионных каналов и передача нервного импульса. Химический синапс
Перенос мембран. Везикулярный транспорт. Эндоцитоз, эндосомы, лизосомы, экзосомы, их особенности. Окаймленные везикулы, COP I, COP II, клатрин. Отшнуровывание везикул. Аппарат Гольджи и его функции как транспортного хаба. Гликозилирование белков в аппарате Гольджи. Участие инозитолфосфатов как маркеров направления транспорта. Rab белки, их участие в адресации везикулярного транспорта. Динамичность адресующих маркеров везикулярного транспорта. Слияние везикул. Возвратный транспорт.
Геномы и их сравнительный размер. Нуклеоид бактерий. Бактериальная транскрипция. Понятие о регуляторных белках. Промоторы и опероны. Транскрипция прокариот: инициация и сигма-факторы, терминация. ДНК в эукариотах. Хромосомные наборы и понятие кариотипа. Расположение хромосом в интерфазном ядре, хромосомные территории. Структура и химические особенности хроматина. Основные белки хроматина - гистоны. Ковалентные модификации гистонов. Минорные гистоны. Нуклеосомы, их строение и структурная роль. Уровни компактизации хроматина. Эухроматин/гетерохроматин, ремоделирование хроматина. Транскрипционно активные домены. Инсуляторы и барьерные белки. Транскрипция эукариот. Сборка преинициаторного комплекса. Медиатор. Энхансеры. Влияние перестройки хроматина. Понятие о транскрипционных факторах.
Cтроение эндоплазматического ретикулума.(ЭР). Основные функции гладкого и шероховатого ЭР. Синтез липидов на примере фосфатидилхолина в гладком ЭР. Общность мембран ЭР и ядерной оболочки. Гладкий и шероховатый ЭР. Биосинтез белка. Аминокислоты, аминоацилирование тРНК. Процессы инициации, элонгации, терминации трансляции. Формирование структуры белка, понятие о шаперонах. Сигналы деградации, деградация белков протеасомой.
Сигналы локализации и транспорт белков. Локализация рибосом и транспорт белков в эндоплазматический ретикулум (ЭР). Встраивание белков в мембрану. Модификации белков в ЭР. Контроль сворачивания белков в ЭР, деградация неправильно свернутых белков, обратная связь от неправильно свёрнутых белков. Транспорт белков в митохондрии, прохождение ими двойной мембраны. Схожесть локализации белков во внешней мембране митохондрий и бактерий. Особенности транспорта и локализация белков в хлоропластах. Транспорт белков в пероксисомы.
Митохондрии: строение и функции. Авторепродукция митохондрий. Геном митохондрий. Элементы биоэнергетики эукариот. Гликолиз. Окислительное фосфорилирование, элементы цикла Кребса. Понятие о B-окислении жирных кислот Строение хлоропластов. Фотосинтез. Хлорофилл, II и I фотосистемы (кратко).
Организация актиновых филаментов (микрофиламентов), промежуточных филаментов, микротрубочек. Принципы сборки актиновых филаментов, актин-ассоциированные белки. Актин и миозины. Цитоскелет прокариот.
Динамика цитоскелета эукариот. Центросомы, центры организации микротрубочек. Микротрубочки и транспорт органелл. Регуляция организации цитоскелета эукариот. Промежуточные филаменты, септины, связь с другими фрагментами цитоскелета.
Репликация ДНК. ДНК-полимераза. Репликативная вилка у бактерий. Сверхспирализация ДНК, хеликаза, топоизомераза. Деление прокариот. Организация перетяжки и распределение генетического материала по дочерним клеткам.
Митоз. S-фаза, биосинтез ДНК, репликация. М-фаза. G1 и G2 фазы. Регуляция клеточного цикла. Чекпоинты и циклины. Изменение концентраций циклинов разного типа в течение клеточного цикла. Чекпоинтные киназы.
Мейоз, его функциональная значимость. Диплоидные и гаплоидные клетки, гаметы. Стадии мейоза, мейоз I и мейоз II. Понятия бивалента и хиазма. Профаза I: стадии; коньюгация и рекомбинация хромосом. Гомологичная рекомбинация, разрешение структур Холлидея. Кроссинговер хромосом: участки кроссинговера и гетеродуплексов. Слияние гамет на примере мыши.
Клеточная гибель: некроз и апоптоз. Программируемая клеточная гибель: апоптоз, его ключевые регуляторы: каспазы, белки группы Bcl2. Внешний и внутренний пути активации апоптоза. Белок р53 и его функционал. Примеры других механизмов клеточной гибели.
Типы рецепторов. G-белок-связанные рецепторы. Передача сигнала через фермент-сопряжённые рецепторы. Рецепторные тирозинкиназы. Понятие о сигнальном каскаде. ГТФаза RAS. Киназные каскады. MAP-киназный каскад. Rho-ГТФазы, цитоскелет. Регуляторные каскады и транскрипционная регуляция. Понятие о каскадах Notch , Wnt, Hedgehog, NFκB. Ядерные рецепторы как пример лиганд-зависимой регуляции.
Введение в межклеточные взаимодействия. Межклеточная адгезия. Кадгерины. Щелевые контакты. Межклеточные переходы, адгезия клеток и внеклеточный матрикс. Интегрины.
Обзор типов клеток человека и особенностей их строения. Клетки крови, мышц, нервной ткани. Стволовые клетки и их особенности. Опухолевые клетки и их признаки.
Базы биологических данных. NCBI: Pubchem, PMC; Blast. Геномные браузеры. Компьютерное представление молекул. Базы данных по биоактивности молекул. Фармакофорные группы.
Устройство микроскопа. Световая микроскопия амёбы: ядро, эндоплазма, эктоплазма, пищевые вакуоли, псевдоподии. Световая микроскопия срезов тканей и клеток клеточных линий млекопитающих. Ядро, клеточная стенка.
Световая микроскопия срезов тканей и клеток клеточных линий млекопитающих. Ядро, клеточная стенка. Органеллы: аппарат Гольджи, митохондрии в растительной и животной клетках. Выявление сайтов транскрипции рибосомной РНК в ядрышках. Световая микроскопия делящихся клеток в тканях и клеточных линиях. Деление клетки, митоз (фазы). Апоптоз в клеточной культуре.
23. Правила работы с бактериальными культурами. Трансформация и работа с клонами бактерий. Пересев и хранение бактериальных клеток, получение музеев бактериальных продуцентов.
Изучение правил работы с бактериальными культурами – устройство ламинарного шкафа, приемы работы в стерильных условиях; приготовление питательных сред и принадлежностей, стерилизация фильтрацией, заливка чашек Петри, хранение бактериальных штаммов (замораживание, столбики агара, чашки), способы рассева (штрихи, шпатель Дригальского) бактериальных клеток из стоков на твердую питательную среду. Фазы роста клеток, титр клеточной культуры и его определение; компетентность бактериальных клеток и трансформация плазмидами, несущими гены флуоресцентных белков (визуальные маркеры экспрессии). Рассев трансформированных клеток на чашки Петри с селективной средой, обсуждение условий роста бактерий на селективной среде. Пересев бактериальных культур. Хранение бактерий (замораживание с глицерином). Рисунок разными экспрессионными штаммами по чашке Петри.
Изучение правил работы с клеточными линиями млекопитающих. Стерильные зоны, особенности работы в стерильных условиях; приготовление питательных сред и растворов высев клеток со стоков. Микроскопический контроль морфологии. Смена среды/пересев клеточных линий. Рассев на планшеты для окрашивания и функциональных тестов. Криоконсервация клеток. Низкотемпературные холодильники и криохранилища.
Прижизненное окрашивание органелл клеток в планшетах флуорофорами (Hoegst или Mitotracker), определение на флуоресцентном микроскопе. Микроскопический контроль плотности посева. Оценка цитотоксичности препаратов. Скретч-тест: зарастание царапины в тесте подвижности. Анализ зависимостей доза-ответ.
26. Эукариоты vs прокариоты. Работа с дрожжевой клеточной культурой, окрашивание дрожжевых и бактериальных клеток.
Сопоставление базовых методов работы с бактериями и дрожжами (жидкая и полутвердая среды, пересевы, колонии). Посев бактериальных культур на чашки с различными антибиотиками. Посев дрожжевых клеток на чашки с теми же антибиотиками. Окрашивание дрожжевых и бактериальных клеток: окрашивание в суспензии, нанесение и фиксация на стекле и просмотр стекол под оптическим микроскопом .
Элементы контроля
Контрольная работа N1
Контрольная работа N2
Экзамен модуля 4
Контрольная работа N3
Экзамен модуля 3
Эндоплазматический ретикулум и митохондрии. Клеточный цикл
Организация клетки. ЭР и аппарат Гольджи
Устройство и функционирование эндоплазматического ретикулума и аппарата Гольджи

Эндоплазматический ретикулум [ЭР (ER)] — протяженная замкнутая мембранная структура, построенная из сообщающихся трубкообразных полостей и мешочков, называемых цистернами. В области ядра ЭР сообщается с внешней ядерной мембраной. Между шероховатым и гладким ЭР имеется морфологическое различие: мембраны шероховатого ЭР усеяны множеством рибосом, в то время как гладкий ЭР не имеет связанных рибосом.
А. Шероховатый эндоплазматический ретикулум и аппарат Гольджи
Шероховатый ЭР [ШЭР (rER)]( 1 ) — место активного биосинтеза белков. Именно здесь синтезируются белки, которые будут функционировать в составе мембран, лизосом или секретироваться из клетки. Остальные белки синтезируются в цитоплазме на рибосомах, не связанных с мембранами ЭР.
Белки, синтезированные на шероховатом ЭР ( 1 ), претерпевают посттрансляционные модификации (созревание белков, см. с. 226). Они либо остаются внутри шероховатого ЭР в виде мембранных белков, либо транспортируются с помощью везикул ( 2 ) в аппарат Гольджи ( 3 ). Транспортные везикулы образуются почкованием мембран, а затем исчезают, сливаясь с ними (см. с. 230).
Подобно ЭР, аппарат Гольджи ( 3 ) представляет собой сложную сеть ограниченных мембранами полостей, имеющих форму диска и являющихся местом созревания и сортировки белков. Имеются цис -, промежуточная и транс -Гольджи-области и транс -Гольджи-сеть . Посттрансляционная модификация белков имеет место в разных областях аппарата Гольджи.
Наконец, созревшие (модифицированные) белки переносятся везикулами в различные отделы клетки, такие, как лизосомы ( 4 ), цитоплазматическая мембрана ( 6 ) или секреторные пузырьки ( 5 ). Последние высвобождают свое содержимое к межклеточное пространство, сливаясь с плазматической мембраной ( экзоцитоз ). Эти транспортные процессы могут быть конститутивными, т.е. проходить постоянно, или регуляторными, т.е. управляться химическими сигналами. Направленность процесса в первую очередь зависит от сигнальной последовательности синтезируемого белка (см. с. 232).
Наряду с белками в аппарате Гольджи осуществляется транспорт мембранных липидов.
Б. Гладкий эндоплазматический ретикулум
ЭР, не имеющий связанных рибосом, называется гладким эндоплазматическим ретикулумом (ГЭР). Он занимает в клетке Сравнительно небольшой объем. Выраженный ГЭР имеется е клетках с активным обменом липидов, таких, как гепатоциты и клетки Лейдига. Для ГЭР характерна замкнутая система разветвленных канальцев.
ГЭР принимает участие в синтезе липидов . Биосинтез осуществляется ферментами, закрепленными на мембранах ГЭР. Здесь локализован синтез фосфолипидов и отдельные стадии синтеза холестерина (см. с. 174). В ГЭР специализированных клеток эндокринной системы протекают различные стадии синтеза стероидных гормонов (см. с. 364). В ГЭР локализованы также процессы метаболической трансформации ксенобиотиков (реакция 1 , см. с. 308). В этих реакциях принимает участие система цитохрома Р450 (см. с. 310), которую считают основной системой ГЭР.
ГЭР выполняет функцию депо ионов Са 2+ , поддерживающего низкий уровень Са 2+ в цитоплазме. Эта функция более всего свойственна саркоплазматическому ретикулуму, специализированной форме ГЭР мышечных клеток (см. с. 326). В мембранах ГЭР локализованы управляемые Са 2+ -каналы и энергозависимые Са 2+ -насосы, а высокая концентрация ионов Са 2+ в цистернах поддерживается при участии Са 2+ -связывающих белков.
Эндоплазматический ретикулум и митохондрии. Клеточный цикл
Эндоплазматический ретикулум и митохондрии. Клеточный цикл
Цитоплазма пронизана сетью мембран, которые образуют энлоплазматический ретикулум (ЭР) и аппарат Гольджи. ЭР представлен непрерывным лабиринтом, состоящим из складчатых мембран, являющихся продолжением наружной ядерной мембраны. ЭР бывает двух типов, которые являются частью одной и той же мембранной системы. На шероховатом ЭР расположены рибосомы — мелкие частицы, на поверхности которых происходит синтез белка, тогда как на гладком ЭР рибосомы отсутствуют.
Аппарат Гольджи представляет собой систему соединенных в виде стопки пузырьков, или цистерн. Белки, подвергшиеся модификации в ЭР, поступают в аппарат Гольджи, где претерпевают дальнейшие модификации и затем покидают его.
Основная функция ЭР и аппарата Гольджи заключается в сортировке белков в зависимости от их предназначения; она осуществляется на основании информации, содержащейся в аминокислотной последовательности белка. F 1роцесс упаковки и направленного транспорта белков к соответствующим участкам клетки или из клетки называют сортингом, или трафиком белков. Митохондрии — это находящиеся в цитоплазме специализированные органеллы, которые при окислении углеродсодержащих соединений, таких как сахара и лиииды, генерируют энергию, сохраняемую в виде аденозинтрифосфата (АТФ). Лизосомы представляют собой окруженные мембраной тельца, содержащие гидролитические ферменты, осуществляющие в клетке дальнейшее образование белков и других макромолекул. Форма клетки определяется ее цитоскелетом, который состоит из сети белковых волокон, тянущихся вдоль и поперек клетки.
Белковые волокна представлены белками трех классов: филаментами актина, микротрубочками и промежуточными филаментами.
Важнейшим содержимым ядра является генетический материал. Структура ядра гранулирована благодаря хроматину, который можно легко увидеть с помощью специальных красителей. В период между двумя делениями хроматин образует плотную единую массу. Во время деления клетки можно видеть, что хроматин состоит из отдельных нитеобразных структур, называемых хромосомами. Общим свойством всех клеток, за исключением достигших конечной специализированной стадии — терминальной дифференцировки, является их способность к делению. В процессе деления в клетке происходит целый ряд структурных изменений. Существенной перестройке подвержены мембраны и цитоскелет.

В клетке образуется новая структура, называемая веретеном; его функция состоит в распределении хромосом между дочерними клетками. В ходе этих изменений приостанавливаются предыдущие функции клетки, включая экспрессию генов, синтез и секрецию белков, способность к движению и гибель клетки.
Клеточный цикл (КЦ), т.е. период между появлением образованной в результате деления новой клетки и ее последующим делением на две дочерние клетки, состоит из двух частей. Интерфаза (относительно длинный период) — это время, в течение которого в клетке протекают процессы синтеза и воспроизводства внутриклеточных компонентов. Во время интерфазы клетка содержит дискретное ядро с компактной массой хроматина. В процессе митоза (относительно короткого периода времени) происходит разделение материнской клетки на две дочерние. В ходе митоза веретено замещает собой внутриклеточные структуры, при этом становятся видны отдельные хромосомы.
Клетки, образующиеся в результате серии митотических делений и формирующие организм, называют соматическими. В процессе эмбрионального развития многие соматические клетки, если не большинство, проходят через КЦ.
Во взрослом организме большинство клеток достигают терминальной дифференцировки и больше не делятся. Они остаются в стационарной фазе (когда синтез ДНК не происходит) — состоянии, эквивалентном бесконечно длящейся интерфазе.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Урок 5. Мембранные органоиды клетки. Ядро. Прокариоты и эукариоты
Мембранные органоиды клетки. Ядро. Прокариоты и эукариоты
Необходимо запомнить
Органоиды клетки
Органоиды, или Органеллы, – постоянные специфические структуры цитоплазмы, выполняющие определённые функции, необходимые для поддержания жизнедеятельности клетки.
Различают органоиды общего значения и специальные органоиды. Органоиды общего значения имеются во всех клетках и выполняют общие функции. Это – митохондрии, рибосомы, эндоплазматическая сеть, комплекс Гольджи, лизосомы, пероксисомы, цитоскелет и клеточный центр.
Органоиды специального значения имеются только в клетках какого-то определённого типа и обеспечивают выполнение функций, присущих только этим клеткам.
Эндоплазматическая сеть (ЭПС) открыта К. Портером в 1945 году. ЭПС или ЭПР (эндоплазматический ретикулум) – сеть канальцев и цистерн, сложенных мембранами. Различают гранулярную (шероховатую, зернистую) и гладкую (агранулярную) ЭПС.
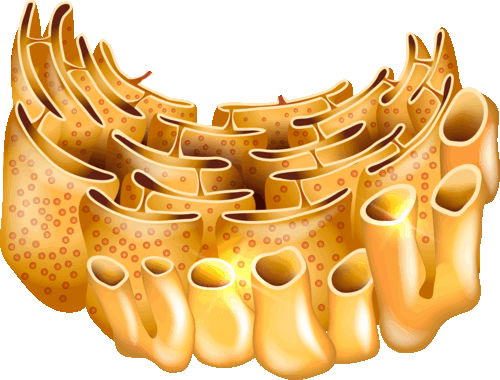
Гранулярная ЭПС содержит рибосомы на наружной стороне мембраны. Гладкая ЭПС не содержит рибосомы. В скелетных мышцах ЭПС носит название саркоплазматический ретикулум. ЭПС пронизывает всю клетку. Полость ЭПС сообщается с ядром и цитоплазматической мембраной.
На рибосомах гранулярной ЭПС синтезируются секреторные белки, предназначенные для выведения из клетки, а также белки лизосом и внеклеточного матрикса.
Наряду с секреторными белками на гранулярной ЭПС синтезируется большая часть полуинтегральных и интегральных белков. В гладеой ЭПС происходит также синтез мембраны липидов и осуществляется «сборка» компонентов мембраны.
Кроме того, ЭПС, как считают, участвует в образовании пероксисом. Таким образом, гранулярная ЭПС служит «фабрикой» мембран для плазмалеммы, аппарата Гольджи, лизосом и других мембранных структур клетки.
Агранулярная (гладкая) эндоплазматическая сеть представляет собой замкнутую сеть трубочек, канальцев, цистерн. На цитоплазматической поверхности гладкой ЭПС синтезируются жирные кислоты, большая часть липидов клетки, в том числе почти все липиды, необходимые для построения клеточных мембран. Поэтому гладкую ЭПС нередко называют «фабрикой липидов». Например, в клетках печени с мембранами гладкого эндоплазматического ретикулума связан фермент, обеспечивающий образование глюкозы из глюкозо-6-фосфата. Эта реакция имеет большое значение в поддержании уровня глюкозы в организме человека.
В организме человека эндоплазматическая сеть особенно хорошо развита в клетках, синтезирующих гормоны, в клетках печени.
Комплекс Гольджи (КГ, или аппарат Гольджи, – пластинчатый комплекс, расположен вблизи ядра, между ЭПС и плазмалеммой. Его структурно-функциональная единица – диктиосома – представляет собой стопку из 5–20 плоских одномембранных мешочков (цистерн), имеющих диаметр около 1 мкм, внутренние полости которых не сообщаются друг с другом. Количество таких мешочков в стопке обычно не превышает 5–20, а расстояние между ними составляет 20–25 нм.
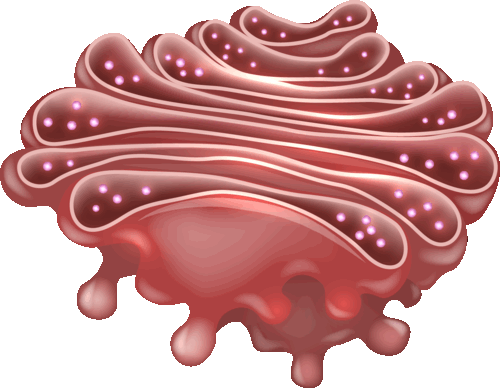
Белки, синтезированные на шероховатой эндоплазматической сети, попадают в аппарат Гольджи. Здесь осуществляется химическая модификация транспортируемых белков и их упаковка в специальные пузырьки.
Таким образом, основными функциями комплекса Гольджи являются химическая модификация, накопление, сортировка, упаковка в секреторные пузырьки и транспорт по назначению белков и липидов, синтезированных в ЭПС.
В комплексе Гольджи образуются лизосомы и синтезируются некоторые полисахариды.
Лизосомальная система и пероксисомы
Лизосомы – мембранные органеллы клеток животных и грибов, содержащие гидролитические ферменты и осуществляющие гидролитическое расщепление макромолекул (внутриклеточное пищеварение). Лизосомы представляют собой окружённые одинарной мембраной пузырьки, размеры которых в клетках животных колеблются от 0,2 до 0,5 мкм. В лизосомах содержится не менее 60 гидролитических ферментов, которые расщепляют все основные классы органических макромолекул.
Все ферменты лизосом активны лишь в кислой среде при значениях pH, близких 5,0. Количество лизосом в разных клетках варьирует от единичных до нескольких сотен, как например, в фагоцитах.
Завершающие этапы процесса внутриклеточного переваривания веществ, поглощённых клеткой, осуществляются в лизосомах.
Лизосомы с помощью своих ферментов могут разрушать не только отдельные органеллы или клетки, но и целые органы (автолиз). Например, в процессе онтогенеза лягушки с помощью ферментов лизосом лизируются хвост и жабры головастика, а образующиеся при этом продукты распада используются для формирования органов взрослого животного.
Митохондрии – крупные мембранные органоиды клетки, которые можно различить в световой микроскоп. Митохондрии присутствуют во всех эукариотических клетках человека, кроме эритроцитов.

Они имеют обычно округлую, удлиненную или нитевидную формы. Количество митохондрий в клетке колеблется в широких пределах (от 1 до 100 тыс. и более) и зависит от потребностей клетки в энергии. Митохондрии имеют наружную и внутреннюю мембраны.
На внутренней поверхности увеличенного фрагмента кристы видны небольшие выпуклости, обращенные в митохондриальный матрикс, которые содержат ферментные системы, обеспечивающие процессы дыхания. Наружная мембрана гладкая и по своему составу сходна с плазмалеммой.
В матриксе содержатся кольцевая молекула митохондриальной ДНК (мтДНК), различные включения, а также молекулы мРНК, транспортной РНК (тРНК) и рибосомы, сходные по строению с рибосомами бактерий. Здесь же располагаются ферменты, превращающие пируват и жирные кислоты в ацетил-КоА, и ферменты реакций цикла Кребса.
Митохондриальная ДНК имеет не линейную, как в хромосомах ядра, а кольцевую форму. Главная функция митохондрий – синтез АТФ, основного источника энергии для обеспечения жизнедеятельности клетки. Поэтому митохондрии называют «энергетическими станциями» клетки.
Пластиды – это органоиды клеток растений и некоторых фотосинтезирующих простейших. У большинства животных и грибов пластид нет.
Пластиды делятся на несколько типов: хлоропласты, хромопласты, лейкопласты. Наиболее важный и известный – хлоропласт, содержащий зелёный пигмент хлорофилл, который обеспечивает процесс фотосинтеза.
Все виды пластид связаны между собой общим происхождением или возможным взаимопревращением. Пластиды развиваются из пропластид – более мелких органоидов меристематических клеток.

Строение пластид
Пластиды относятся к двумембранным органоидам, у них есть внешняя и внутренняя мембраны.
Во многих пластидах, особенно в хлоропластах, хорошо развита внутренняя мембранная система, формирующая такие структуры, как тилакоиды, граны (стопки тилакоидов), ламелы – удлинённые тилакоиды, соединяющие соседние граны. Внутреннее содержимое пластид обычно называют стромой. В ней, помимо прочего, находятся крахмальные зёрна.
Считается, что в процессе эволюции пластиды появились аналогично митохондриям – путём внедрения в клетку-хозяина другой прокариотической клетки, способной в данном случае к фотосинтезу. Поэтому пластиды считают полуавтономными органеллами. Они могут делиться независимо от делений клетки, у них есть собственная ДНК, РНК, рибосомы прокариотического типа, т. е. собственный белоксинтезирующий аппарат. Часть генов, управляющая их функционированием, находится как раз в ядре.
Ядро – важнейшая часть эукариотической клетки. Оно состоит из ядерной оболочки, кариоплазмы, ядрышек, хроматина.
1. Ядерная оболочка по строению аналогична клеточной мембране, содержит поры. Ядерная оболочка защищает генетический аппарат от воздействия веществ цитоплазмы. Осуществляет контроль за транспортом веществ.
2. Кариоплазма представляет собой коллоидный раствор, содержащий белки, углеводы, соли, другие органические и неорганические вещества. В кариоплазме содержатся все нуклеиновые кислоты: практически весь запас ДНК, информационные, транспортные и рибосомальные РНК.
3. Ядрышко – сферическое образование, содержит различные белки, нуклеопротеиды, липопротеиды, фосфопротеиды. Функция ядрышек – синтез зародышей рибосом.
4. Хроматин (хромосомы). В стационарном состоянии (время между делениями) ДНК равномерно распределены в кариоплазме в виде хроматина. При делении хроматин преобразуется в хромосомы.
Функции ядра: в ядре сосредоточена информация о наследственных признаках организма (информационная функция); хромосомы передают признаки организма от родителей к потомкам (функция наследования); ядро согласует и регулирует процессы в клетке (функция регуляции).
Клеточные структуры

По строению и организации клетки — очень сложные образования (рис. 1). В них обнаружены различные микроскопические и субмикроскопическне структуры, обладающие высокой динамичностью и способностью закономерно изменяться соответственно изменениям условий существования организма и приспосабливаться к ним.
Наиболее важные компоненты клетки (животной и растительной)— ядро и цитоплазма, которые тесно связаны между собой и не могут существовать раздельно, однако строение и функции их неодинаковы. Ядро возникло на определенном этапе эволюционного развития клетки.
Ядро — главный органоид клетки. Оно покрыто тонкой двухслойной мембраной с порами для соединения с цитоплазмой. Ядро обычно имеет шарообразную форму (рис. 2), но в некоторых случаях — палочкообразную, лопастную и даже сетчатую. В ядре хорошо видна еще меньшая по размеру структура — ядрышко, где накапливается и, по-видимому, синтезируется РНК, которая затем переносится в цитоплазму и становится основной структурной единицей рибосом, В ядрах содержится большое количество ферментов, обусловливающих освобождение и трансформацию энергии и осуществление многочисленных синтезов. Таким образом, энергетически ядра представляют собой полностью независимые органоиды клетки. Состав их ферментов обеспечивает образование аденозинтрифосфорной кислоты (АТФ) в процессе гликолиза и свидетельствует о высокой метаболической активности.
Химическими компонентами ядра являются в основном: белки и нуклеиновые кислоты. Химический состав изолированных ядер, выделенных из проростков гороха (по А. Фрей-Висслингу и К. Мюлеталеру), % (по массе): ДНК—14, РНК—12,1, основные белки — 22,6, другие белки — 51,4.
Основные (щелочные) белки — это протамины и гистоны, которые содержат много основных аминокислот: лизина, гистидина и особенно аргинина. Протамины и гистоны образуют с нуклеиновыми кислотами нуклеопротеиды. Другие белки могут содержаться в ядре в виде самостоятельной фазы. Ядрышко состоит из больших гранул, которые по размеру близки к рибосомам (диаметр их 15 нм) и содержат большое количество РНК. Основное вещество ядра называется нуклео-плазмой, В ядре находятся хромосомы — носители наследственности, Хромосомы имеют хроматиновые структуры, основными компонентами которых являются ДНК и РНК.
Цитоплазма — рабочий аппарат клетки. Она пронизана системой мембран, которые отходят от ядерной оболочки и соединяются с внешней мембраной клетки. Эти внутренние клеточные мембраны, образующие густо переплетенную сеть с многочисленными канальцами и полостями, называют эндоплазматической сетью. Функциональное значение ее разнообразно, Она участвует в синтезе ассимилятов. Мембраны эндоплазматической сети пронизывают и объединяют в единое целое клетку, а также служат своеобразными путями перемещения ассимилятов и передачи раздражения от клетки к клетке, которая осуществляется последовательной перезарядкой поверхности мембран. Химически клеточные мембраны гетерогенны и состоят из протеидов (80%, из них 25% приходится на белки-ферменты) и липидов (20%), среди которых много фосфолипидов.
Структурные элементы клетки можно разделить на три большие функциональные группы:
1) органеллы, которые катализируют превращение энергии, — митохондрии и хлоропласта;
2) органеллы, которые катализируют репликацию белков, — рибосомы, полирибосомы;
3) клеточные гранулы и другие образования, которые принимают участие в синтетических реакциях, обмене веществ (сферосомы, цитосомы, элайопласты, транслосомы, осмиофильные гранулы, аппарат Гольджи и т. д.).
Электронно-микроскопическими исследованиями установлено, что растительная клетка состоит из большого количества мембранных систем. Многие из субклеточных компонентов имеют мембранное строение.
В процессе эволюции хондриом (совокупность митохондрий клетки) образовался, по-видимому, позже, чем пластом (совокупность пластид — лейкопласты, хлоропласта и хромопласты). Возможно, что митохондрии отделились в филогенезе от системы пластид. Митохондриальный аппарат — необходимый компонент всех клеток, которым присущ аэробный способ жизни, тогда как хлоропласты являются обязательным ингредиентом только клеток автотрофных растений.
Хлоропласты и митохондрии — это «силовые станции», от которых зависит жизнедеятельность клетки и растения в целом, так как одна из них накапливает солнечную энергию в макроэргических связях АТФ, а вторая превращает энергию, содержащуюся в питательных веществах, также в энергию АТФ.
Митохондрии (от греч. mitos — нить, chondros — зернышко) — органеллы шарообразной формы, диаметром 0,5 мкм и длиной 2 мкм. Это нестойкие структуры, в липофильных жидкостях они разрушаются, а в воде набухают, имеют двойную оболочку, состоящую из внутренней и внешней мембран. Между мембранами имеется просвет (10 нм), заполненный сывороткой. Внутренняя мембрана митохондрий образует кристы, или трубочки. К ней со стороны стромы присоединены с помощью «стебельков», или ножек, частицы, которые получили название оксисом, так как они содержат окислительные ферменты. Внутреннее пространство митохондрий заполнено матриксом, или хондриоплазмой, — вязким раствором, содержащим ферменты (рис. 3, 4).
Митохондрии состоят из белка (2/3) и липидов (1/3), среди которых половина приходится на фосфолипиды. Неотъемлемыми компонентами митохондрий являются ДНК и все типы РНК. Находящаяся в матриксе митохондрий ДНК в виде нитей способна к независимой от ДНК ядра репликации. В митохондриях обнаружены специфические рибосомы, которые обеспечивают автономный синтез некоторых белков. Так, митохондрии проростков гороха содержат, %: белка — 30-40, РНК — 0,5-1, фосфолипидов — 30. В митохондриях сосредоточены ферменты цикла трикарбоновых кислот, флавопротеиды и цитохромы.
Митохондрии — дыхательные центры клетки, обладают следующими функциями: 1) осуществляют окислительные реакции, являющиеся источником электронов; 2) переносят электроны по цепи компонентов, синтезирующих АТФ; 3) катализируют синтетические реакции, идущие с использованием энергии АТФ; 4) регулируют биохимические процессы в цитоплазме.
При анаэробном дыхании митохондрии исчезают и эндоплазматическая сеть становится более развитой. В клетке в зависимости от ее типа и функции содержится 50—5000 митохондрий.
Пластиды образуются из пропластид — маленьких амебообразных телец (диаметром 0,05—0,5 мкм), которые формируются из инициальных частиц, отделяющихся от ядра, и содержат нуклеоплазму. Пластом состоит из пластид трех типов: лейкопластов (бесцветные), хлоропластов (зеленые) и хромопластов (оранжевые); их размеры в среднем составляют 2×5 мкм. Пластиды по монотропному типу развития проходят несколько стадий, а именно (по А. Фрей-Висслингу и К. Мюлеталеру);
| 1) ювенильная (рост) | инициальные частицы протопластиды |
| 2) активная (метаболизм) | лейкопласты хлоропласты |
| 3) дегенерация, старение (пассивное состояние) | хромопласты |
Пластиды имеют двойную мембрану, внутри которой находится гранулярное вещество, называемое стромой. В начале развития строма пластид имеет гранулярное строение.
Структура хлоропласта формируется в несколько этапов. Первичная дифференциация пластиды начинается с инвагинаций внутренней мембраны до образования проламеллярного тела (без света). Второй этап связан с образованием ламеллярно-гранулярного строения, биосинтезом и накоплением хлорофилла. В хлоропластах высших растений образуются граны, которые состоят из серии ламелл, или двойных мембран (рис. 5). Каждая двойная мембрана образует закрытый мешочек, или сумку, которая называется тилакоидом. Ламеллы состоят из белков и липидов. Химический анализ ламелл, выделенных из хлоропластов шпината, показал, что в них 52% приходится на белок и 48% на липидную фракцию, которая включает хлорофилл а и b, каротиноиды (ксантофиллы к каротины), пластохинон, витамин К1 фосфолипиды (галактозил-глицериды, фосфоглицериды), сульфолипиды.
Хлоропласты имеют определенную биохимическую и генетическую автономность. В них синтезируется ДНК, которая отличается от ядерной ДНК. Хлоропластам свойственны также своя белоксинтезирующая система (рибосомы) и автономность процесса биосинтеза белка. Увеличение размеров пластид коррелирует с накоплением белка. Хлоропласт можно представить как уменьшенную и упрощенную модель клетки, которая реагирует на .действие света включением и выключением своих генов. Основная функция хлоропластов — участие в процессе фотосинтеза. Функция лейкопластов — участие во вторичном синтезе крахмала в клетках. Хромопласты, как правило, образуются из хлоропластов, а иногда, например в корнеплоде моркови, из лейкопластов. При этом строма дегенерирует, ламеллярная структура разрушается, оболочка сохраняется и образуются глобулы, которые содержат капельки желтого цвета с каротиноидами.
Рибосомы — это рибонуклеопротендные частицы сферической формы, диаметром 15—35 нм. В них содержится приблизительно одинаковое количество структурного белка и высокополимерной РНК. Комплексы из пяти и более рибосом называются полирибосомами, или полисомами. Каждая рибосома состоит из двух субъединиц с различными коэффициентами седиментации, которые агрегатируются в единое целое с помощью ионов магния. Слипаясь по две, они образуют димеры. Рибосомы очень пористые и отличаются высокой степенью гидратации. Они выполняют чрезвычайно важные функции в обмене веществ—это центры биосинтеза белка в клетке. Роль рибосом в белковом синтезе заключается в том, что они обусловливают процесс, при котором активированные аминокислоты конденсируются, образуя полипептидную цепь.
Рибосомы имеются в клетках всех организмов. Они локализуются в цитоплазме и органеллах (ядре, митохондриях, пластидах). В хлоропластах рибосомы находятся в свободном и связанном с их структурой состояниях. Последние более интенсивно включают 14 С аминокислот в белки и являются полирибосомами. В дифференцированных клетках большинство рибосом, как правило, связаны с липопротеидными мембранами, которые, пронизывают цитоплазму и образуют эндоплазматическую сеть. Такая эндоплазматическая сеть с прикрепленными к ней рибосомами называется эндоплазматическим ретикулумом.
Сферосомы — субмикроскопические компактные частицы цитоплазмы диаметром 0,4—0,8 мкм, содержат белковую строму и цитохромоксидазу, ферментативно активны, богаты жирами. Сферосомы, по-видимому, осуществляют биосинтез жиров, а именно последний его этап — переэтерификацию глицерофосфата путем обмена между фосфорной кислотой и жирными кислотами. Таким образом, сферосомы можно считать специализированными органеллами, функция которых — биосинтез жиров.
Лизосомы по структуре и химическому составу близки к сферосомам, но богаче ферментами — в них обнаружены ферменты нуклеазы, фосфатазы, протеазы и т.д. Вполне вероятно, что лизосомы переваривают макромолекулярные продукты, поглощенные путем пиноцитоза. Они участвуют и в автолизе клетки.
Цитосомы — мелкие гранулы, которые находятся в тесном контакте с мембранами эндоплазматической сети, чем и отличаются от свободнолежащих сферосом и лизосом.
Транслосомы — толстостенные гранулы, функция которых, по-видимому, заключается в накоплении продуктов метаболизма фенольных производных и их транспортировке в вакуоль.
Так называемые диктиосомы, или аппарат Гольджи, могут состоять из отдельных пластинок, палочек и чешуек, разбросанных по всей цитоплазме клетки. Возможно, что аппарат Гольджи принимает участие в управлении общим ходом физиологических процессов, в, образовании вакуолей и клеточных оболочек.
Таким образом, высокая активность органелл клетки связана с особенностями их строения и химическим составом. Пластиды, митохондрии, микросомы содержат основную массу биокатализаторов (ферментов, витаминов и т.д.) и большое количество (25—30%) липонуклеопротеидов (табл. 2).
| Органеллы | Белки | Липиды | Нуклеиновые кислоты |
| Цитоплазма | 80—95 | 2—3 | 1—2 |
| Пластиды | 30—45 | 20—40 | 0,5—3 |
| Митохондрии | 30—40 | 25—38 | 1—6 |
| Рибосомы | 50—57 | 3—4 | 35 |
| Ядро | 50—80 | 8—40 | 10—30 |
Каждая группа органелл выполняет свои функции на основании непрерывного взаимодействия с другими физиологически активными центрами клеток. Это видно на примере фотосинтеза и дыхания.
Клеточная оболочка состоит из клетчатки, или целлюлозы (C6H10O5)n, — полисахарида, который гидролизуется до глюкозы. Клетчатка является главным веществом хлорофиллоносных растений и по абсолютному количеству занимает первое место среди всех органических веществ на земной поверхности. Ниже приведен химический состав первичных растительных клеточных оболочек (колеоптилей, стеблей, листьев, корневых волосков), % на сухое вещество:
| гемицеллюлоза | 53 |
| целлюлоза | 30 |
| пектиновые вещества | 5 |
| белки | 5 |
| липиды | 7 |
Клеточная оболочка начинает развиваться с образования клеточной пластинки в анафазе митоза. Это происходит сразу после деления ядра. Целлюлоза в виде микрофибрилл формирует каркас. Микрофибриллы — эластичный строительный элемент клеточной оболочки (стенки), диаметр их 10—30 нм, длина несколько микрометров.
Отдельная микрофибрилла состоит из нескольких сотен молекул целлюлозы. Микрофибриллы, располагаясь параллельно своей оси, удерживаются вместе водородными связями и у высших растений цементируются гемицеллюлозой. Характерная особенность микрофибрилл целлюлозы — анизотропия набухания в воде, т.е. не во всех направлениях их поперечного сечения микрофибриллы увеличиваются одинаково. Важнейшая функция клеточной оболочки заключается в физическом противодействии осмотическому давлению со стороны внутреннего содержимого клетки. Это давление является движущей силой увеличения размеров и изменения формы клеток.
Клетчатка составляет более 50% древесины, а в волокне хлопчатника — более 90%. Структурная формула молекулы целлюлозы имеет такой вид:

Молекула клетчатки в среднем содержит 1`400—10`000 глюкозных остатков, расположенных в виде цепочки и соединенных между собой кислородным мостиком. Количество остатков глюкозы в молекуле целлюлозы в зависимости от растительного материала и способа его обработки изменяется в широких пределах (табл. 3). Соответственно изменяется молекулярная масса, или степень полимеризации. В длинных волокнообразных клетках, например в волосках семян хлопчатника или в лубяных волокнах льна, конопли, цепочки целлюлозы вытянуты в одном направлении, по длине клеток, но под некоторым углом к продольной оси. Таким образом, клеточная стенка имеет несколько спиральное строение.
| Растительный материал | Молекулярная масса | Количество остатков глюкозы в молекуле |
| Хлопчатник | 1`750`000 | 10`800 |
| Лен | 5`900`000 | 36`000 |
| Рами | 20`000 | 12`400 |
| Древесина | 400`000—500`000 | 2`500-3`100 |
Между мицеллами целлюлозы остаются межмицеллярные пространства, сквозь которые может легко проходить вода как в середину клетки, так и в окружающую среду. По бокам цепи целлюлозы связаны друг с другом дополнительными валентностями. Последние в несколько раз слабее, чем связи основных валентностей, поэтому волокнообразные клетки сравнительно легко расщепляются в продольном направлении, тогда как в поперечном направлении они достаточно прочные (не уступают в этом отношении металлической проволоке).
В паренхимных клетках цепочковидные молекулы целлюлозы состоят из мицелл, которые перекрещиваются в различных направлениях и образуют волокнообразную структуру или сложную неправильную сетку. Пространства такой сетки заполнены водой и межмицеллярными веществами. В клетках молодых эмбриональных тканей таким межмицеллярным веществом является пектин, а в клетках древесины — лигнин.
Клеточные оболочки способны к набуханию. Около нитевидных молекул целлюлозы вода размещается в продольных щелях, и оболочка набухает главным образом в поперечном направлении и очень слабо в продольном. Иная картина в паренхимных клетках, где наблюдается переплетение мицелл в виде войлока. В этом случае клетки паренхимы, набухая, увеличиваются в объеме во всех направлениях.
Вакуоль. Основная функция вакуоли — поддержание гомеостаза клетки. В клеточном соке вакуоли в растворенном состоянии содержатся соли, сахаристые вещества, белки, аминокислоты, органические кислоты, липиды, а также пигменты, которые относятся главным образом к группе флавоноидов. Так, пигменты антоцианы придают лепесткам цветков и другим частям растения красную, фиолетовую, синюю окраску. Красная окраска корней столовой свеклы обусловливается присутствием в клеточном соке бетанина — гликозида β-цианина (азотсодержащего аналога антоцианина).
Исследования выделенных вакуолей (Калифорнийский университет США) показали, например, что вакуоли из эндосперма прорастающих семян клещевины содержат до 25% всего количества белка в клетке, 62% сахарозы и различные гидролитические ферменты: кислую протеазу, карбопептидазу, фосфодиэстеразу, β-галактозидазу и др.
Получены интересные данные об образовании вакуолей. В клетках сухих семян вакуоли отсутствуют, но в них содержатся белковые тела — отложения запасных белков. При прорастании семян наружный слой белковых тел растворяется в воде и образуются капельки, содержащие их остатки. Затем капельки сливаются, образуя большую центральную вакуоль. Известны также данные о протеолитической функции вакуолей.
Следовательно, опровергается представление о вакуоли как о вместилище конечных продуктов обмена, отходов клетки, не имеющих влияния на биохимические процессы, происходящие в цитоплазме, и вакуоль рассматривается как органелла, обладающая аутофаговой активностью, участвующая в обмене веществ клетки.
Читайте также: