Сбалансированные перестройки хромосом: инверсия
Обновлено: 18.04.2024
Преимплантационное генетическое тестирование хромосомных перестроек — это одно из разновидностей преимплантационного генетического скрининга эмбрионов на наличие/отсутствие хромосомного дисбаланса в их геноме.
Помимо численных аномалий (трисомии, моносомии, полисомии, триплоидии, др.) хромосомный дисбаланс у эмбриона может быть представлен различными структурными аномалиями в самих хромосомах – хромосомные аберрации.
В основе таких хромосомных аномалий лежит первоначальное нарушение целостности хромосомы — разрывы. Повреждение ДНК в виде двухцепочечных разрывов может быть обусловлено внешними факторами (химиотерапией, ионизирующим излучением), структурные перестройки могут наследоваться от отца или матери.
В результате таких разрывов может произойти либо утрата генетического материала (делеция), либо удвоение участка хромосомы – дупликация, либо различные внутрихромосомные и межхромосомные перестройки.
Патогенное влияние структурных дефектов хромосом у эмбриона на исход беременности и на здоровье потомства зависит от типа хромосомной перестройки, её локализации на хромосоме, дозы генов, попавших в участок дефекта/аберрации. Урон, наносимый такими структурными хромосомными деффектами, выразится либо в прекращении эмбриогенеза (развития эмбриона), либо в рождении ребенка с патологическим синдромом.
Структурные хромосомные аномалии в виде делеций, дупликаций и транслокаций встречаются наиболее часто среди иных возможных структурных дефектов и признаны наиболее клинически значимыми.
Делеции – это потеря участка хромосом, потеря генов.
Дупликации – это, напротив, увеличение количества хромосомного материала.
Эти дефекты могут быть разных размеров - от крупных, доступных современным микроскопам, до микроделеций и микродупликаций, которые могут проявляться с достаточной частотой, но которые можно обнаружить только современными молекулярными методами с высоким разрешением (способностью их увидеть). Примеры наиболее частых врожденных синдромов после таких «поломок»: Ди Джорджи, Прадера-Вилли, Ангельмана, «кошачьего крика». Все они сопровождаются не только физическими расстройствами, но и умственной отсталостью.
Транслокации – ещё один из не менее клинически значимых дефектов хромосом. Это межхромосомная перестройка, когда происходит перенос участка из одной хромосомы на другую, негомологичную. Носительство транслокаций встречается практически у каждого из 500 человек, но лишь в 5,5% случаев носительства сбалансированных хромосомных перестроек, по данным наблюдений специалистов, не влияет на реализацию репродуктивной функции.
Особое опасение перед переносом эмбриона в полость матки вызывает риск формирования несбалансированной транслокации у эмбриона, когда родители являются скрытыми носителями сбалансированных перестроек. Сбалансированные перестройки у будущих родителей могут никак не проявлять себя. Проблема всплывает при зачатии, когда сбалансированная перестройка может явиться причиной формирования несбалансированной у эмбриона. Беременность эмбрионом с такой поломкой может закончиться самопроизвольным абортом или привести к появлению ребенка с врожденными дефектами и/или умственной отсталостью.
Обнаружить за одно исследование максимально все возможные такие «поломки» в хромосомном материале эмбрионов могут полногеномные тесты на основе двух современных технологий последнего поколения - aCGH и NGS, которые мы и используем в нашей лаборатории.
Сбалансированные перестройки хромосом: инверсия
Сбалансированные перестройки хромосом: инверсия
Хромосомные перестройки обычно не имеют фенотипического эффекта, если они сбалансированы, поскольку весь хромосомный материал присутствует, даже если он иначе упакован. Важно различать разницу между истинно сбалансированными перестройками и теми, которые цитогенетически представляются сбалансированными, но на молекулярном уровне не сбалансированы.
Более того, из-за высокой частоты в геноме CNP, увеличивающей и без того существующие различия в много миллионов пар оснований между геномами неродственных индивидуумов, понятие сбалансированности или несбалансированности отчасти произвольно и подлежит дальнейшему исследованию и уточнению.
Даже когда структурные перестройки истинно сбалансированы, они могут представлять угрозу последующим поколениям, поскольку носители могут иметь высокую частоту несбалансированных гамет и, следовательно, имеют повышенный риск аномального потомства с несбалансированными кариотипами; в зависимости от специфической перестройки, риск может колебаться от 1 до 20%.
Существует также возможность, что один из хромосомных разрывов повредит ген, вызвав мутацию. Это хорошо описаная причина Х-сцепленных болезней у женщин-носителей сбалансированных транслокаций Х-хромосомы и аутосом, такие транслокации могут быть путеводной нитью к уточнению позиции гена, ответственного за развитие болезни.

Инверсия хромосом
Инверсия появляется, когда в хромосоме происходят два разрыва, а сегмент между ними переворачивается. Инверсии бывают двух типов: парацентрические (не включающие центромеру), при которых оба разрыва происходят в одном плече; и перицентрические (включающие центромеру), при которых разрывы находятся в разных плечах. Поскольку парацентрические инверсии не изменяют соотношение плеч хромосомы, их можно выявить (если это вообще удастся) только при дифференциальной окраске или FISH-методом с локусспецифическими зондами.
Перицентрические инверсии цитогенетически выявить легче, поскольку они могут изменять соотношение плеч хромосом, а также расположение полос.
Инверсия обычно не вызывает аномалий фенотипа у носителей, поскольку относится к сбалансированным перестройкам. Ее медицинское значение — влияние на потомство; носитель любой инверсии попадает в группу риска по наличию аномальных гамет, приводящих к несбалансированному потомству, так как при инверсии при конъюгации хромосом в мейозе I формируется петля. Рекомбинация отчасти подавляется в пределах инверсионных петель, но если она происходит, то может вести к формированию несбалансированных гамет.
Формируются как гаметы со сбалансированным хромосомным набором (нормальным или с инверсией), так и гаметы с несбалансированными хромосомами в зависимости от позиции точек рекомбинации. Если инверсия парацентрическая, несбалансированные рекомбинантные хромосомы обычно ацентрические или дицентрические и не могут привести к жизнеспособному потомству, хотя есть и редкие исключения. Таким образом, на самом деле риск того, что носитель парацентрической инверсии будет иметь живорожденного ребенка с аномальным кариотипом, очень низкий.
Перицентрическая инверсия, со своей стороны, может привести к появлению несбалансированных гамет как с дупликацией, так и с делецией хромосомных сегментов. Удвоенный и потерянный сегменты — расположенные дистальнее инверсии. В целом риск для носителя перицентрической инверсии родить ребенка с несбалансированным кариотипом оценивают в 5-10%. Каждая перицентрическая инверсия, тем не менее, связана с конкретным риском.
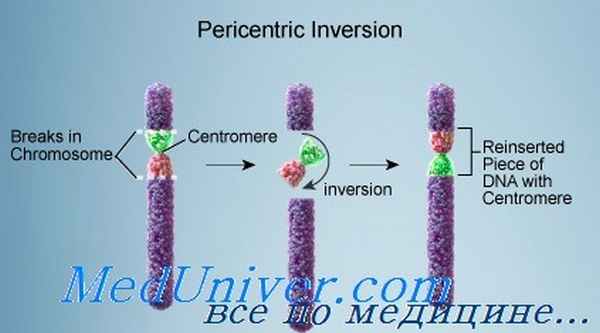
Большие перицентрические инверсии с большей вероятностью, чем малые, приводят к жизнеспособному рекомбинантному потомству, так как в случае больших инверсий несбалансированные сегменты в рекомбинантном потомстве имеют меньший размер. Это положение иллюстрируют три хорошо изученных инверсии.
Перицентрическая инверсия хромосомы 3, происходящая от семейной пары из Ньюфаундленда, вступивших в брак в начале XIX в. — одна из немногих, для которой получено достаточно данных, чтобы оценить расхождение инвертированной хромосомы в потомстве носителей. Кариотип с inv(3) (p25q21) с тех пор был описан в ряде центров Северной Америки, в семьях, предки которых прослеживались до приморских провинций Канады.
Другая перицентрическая инверсия, связанная с выраженным дупликационным или делеционным синдромом в рекомбинантных потомках, находится в хромосоме 8, inv(8) (p23.1q22.1) и первоначально обнаружена среди испанцев юго-запада США. Эмпирические исследования показали, что носители inv(8) имеют 6% риска родить ребенка с синдромом рекомбинантной хромосомы 8, летальным нарушением с серьезными сердечными аномалиями и умственным недоразвитием. В рекомбинантной хромосоме дублирован дистальный участок 8q22.1 и утерян дистальный участок 8р23.1.
Наиболее частая инверсия у человека — небольшая перицентрическая инверсия хромосомы 9, которую отмечают почти у 1% обследованных лабораториями цитогенетики. Кариотип inv(9)(pllql2) не имеет известных опасных эффектов у носителей и не дает значимого риска выкидыша или несбалансированного потомства; поэтому обычно считается вариантом нормы.
Дополнительно к цитогенетически видимым инверсиям геномными методами обнаруживают все большее количество малых инверсий. Полагают, что многие из них клинически благоприятны, без отрицательного влияния на потомство.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
XI Международная студенческая научная конференция Студенческий научный форум - 2019

Типы хромосомных аберраций в эукариотической клетке.
Текст работы размещён без изображений и формул.
Полная версия работы доступна во вкладке "Файлы работы" в формате PDF
Хромосомные аберрации (хромосомные мутации, хромосомные перестройки) — изменения структуры хромосом. Классифицируют делеции (удаление участка хромосомы), инверсии (изменение порядка генов участка хромосомы на обратный), дупликации (повторение участка хромосомы), транслокации (перенос участка хромосомы на другую). Хромосомные перестройки носят, как правило, патологический характер и нередко приводят к гибели организма. Показано значение хромосомных перестроек в видообразовании и эволюции.
ВОЗНИКНОВЕНИЕ ХРОМОСОМНЫХ АББЕРАЦИЙ
В ходе кроссинговера образуются разрывы хромосом, которые затем репарируются. Нарушения процесса репарации могут привести к появлению хромосомных перестроек. Разрывы хромосом и, как следствие, образование перестроек происходят под действием различных мутагенных факторов: физической (ионизирующее излучение), химической или биологической (транспозоны, вирусы) природы. Также некоторые хромосомные перестройки (делеции) характерны для носителей специфических сайтов ломкости.
Различают терминальные (утрата концевого участка хромосомы) и интеркалярные (утрата участка на внутреннем участке хромосомы) делеции. Если после образования делеции хромосома сохранила центромеру, она аналогично другим хромосомам передается при делении, участки же без центромеры как правило утрачиваются. При конъюгации гомологов во время кроссинговера у нормальной хромосомы на месте делеции в мутировавшей хромосоме образуется т. н. делеционная петля, которая компенсирует отсутствие делетированного участка.
Исследованные делеции редко захватывает протяженные участки хромосом, обычно такие аберрации летальны. Самым хорошо изученным заболеванием, обусловленным делецией, является синдром кошачьего крика, описанный в 1963 году Джеромом Леженом. В его основе лежит делеция небольшого участка короткого плеча 5 хромосомы. Для больных характерен ряд отклонений от нормы: нарушение функций сердечно-сосудистой, пищеварительной систем, недоразвитие гортани (с характерным криком, напоминающим кошачье мяуканье), общее отставание развития, умственная отсталость, лунообразное лицо с широко расставленными глазами. Синдром встречается у 1 новорожденного из 50000.
Другой интересной делецией является делеция в гене, кодирующем рецептор CCR5. Этот рецептор используется вирусом иммунодефицита человека (ВИЧ) для распознавания своей цели — Т-лимфоцитов. Продукта гена с делецией получил название CCR5-Δ32, этот вариант CCR5 не узнается ВИЧ, и носители такой мутации к ВИЧ невосприимчивы (это порядка 10 % европейцев).
Дупликации появляются в результате неравного кроссинговера (в этом случае второй гомолог несет делецию) или в результате ошибки в ходе репликации. При конъюгации хромосомы с дупликацией и нормальной хромосомы как и при делеции формируется компенсационная петля.
Практически у всех организмов в норме наблюдается множественность генов, кодирующих рРНК (рибосомальную РНК). Это явление назвали избыточностью генов. Так у E. coli на рДНК (ДНК, кодирующее рРНК) приходится 0,4 % всего генома, что соответствует 5-10 копиям рибасомальных генов.
Другой пример дупликации — мутация Bar у Drosophila, обнаруженная в 20-х годах XX века Т. Морганом и А. Стертевантом. Мутация обусловлена дупликацией локуса 57.0 X-хромосомы. У нормальных самок (B+/B+) глаз имеет 800 фасеток, у гетерозиготных самок (B+/B) глаз имеет 350 фасеток, у гомозигот по мутации (B/B) — всего 70 фасеток. Обнаружены также самки с трижды повторенным геном — double Bar (BD/B+).
В 1970 году Сусумо Оно в монографии «Эволюция путем дупликации генов» разработал гипотезу об эволюционной роли дупликаций, поставляющих новые гены, не затрагивая при этом функций исходных генов. В пользу этой идеи говорит близость ряда генов по нуклеотидному составу, кодирующих разные продукты. Это трипсин и хемотрипсин, гемоглобин и миоглобин и ряд других белков.
Различают парацентрические (инвертированный фрагмент лежит по одну сторону от центромеры) и перицентрические (инвертированный фрагмент лежит по разные стороны от центромеры) инверсии. При инверсиях не происходит потери генетического материала, потому как таковые инверсии как правило не влияют на фенотип, но если в инверсионной гетерозиготе (то есть организме, несущем как нормальную хромосому, так и хромосому с инверсией) происходит кроссинговер, то существует вероятность формирования аномальных хроматид. В случае парацентрической инверсии образуется одна нормальная и одна инвертированная (фенотипически нормальная) хроматиды, дицентрическая хроматида с дупликацией и делецией (при расхождении хроматид она обычно разрывается на две) и ацентрическая хроматида с дупликацией и делецией (обычно утрачивается). В случае перицентрической инверсии образуется одна нормальная и одна инвертированная хроматиды, а также две хроматиды с дупликацией и делецией. Гаметы, несущие дефектные хромосомы, обычно не развиваются или погибают на ранних этапах эмбриогенеза. Но гаметы с инвертированной хромосомой развиваются в организмы, 50 % гамет которых нежизнеспособны. Т.о. мутация сохраняется в популяции.
У человека наиболее распространенной является инверсия в 9 хромосоме, не вредящая носителю, хотя существуют данные, что у женщин с этой мутацией существует 30 % вероятность выкидыша.
Существует несколько форм транслокации:
• собственно транслокация (перенос участка с одной негомологичной хромосомы на другую);
• реципрокная транслокация (две негомологичные хромосомы обмениваются участками);
• робертсоновская транслокация (две негомологичные хромосомы объединяются в одну);
• транспозиция (перенос участка хромосомы на другое место на той же хромосоме).
Транслокация, реципрокная транслокация и транспозиция, которые не сопровождаются утратой генетического материала (т. н. сбалансированные транслокации), часто не проявляются фенотипически. Однако, как и в случае с инверсиями, в процессе гаметогенеза часть сформированных гамет несет летальные аберрации. К примеру, в случае реципрокной транслокации обычно выживает не более 50 % зигот.
Примером транслокации может служить т. н. семейный синдром Дауна. При этом заболевании у одного из родителей обнаруживается фенотипически непроявляющаяся транслокация 21 хромосомы на 14. У такого человека с вероятностью в 1/4 образуются гаметы с двумя 21 хромосомами (одна свободная и одна траслоцированная). При слиянии такой гаметы с нормальной образуется трисомик по 21 хромосоме.
Другой пример — транслокация типа Philadelphia, транслокация между 9 и 22 хромосомами. В 95 % случаев эта мутация является причиной одной из форм хронической лейкемии (chronic myelogenous leukemia).
Робертсоновские транслокации, возможно, являются причиной различий между числом хромосом у близкородственных видов. Существуют данные, что два плеча 2-й хромосомы человека соответствуют 12 и 13 хромосомам шимпанзе. Возможно, 2-я хромосома образовалась в результате робертсоновской транслокации двух хромосом обезьяноподобного предка человека. Таким же образом объясняют тот факт, что различные виды дрозофилы имеют от 3 до 6 хромосом.
Робертсоновские транслокации привели к появлению в Европе нескольких видов-двойников (хромосомные расы) у мышей группы видов Mus musculus, которые, как правило, географически изолированы друг от друга. Набор и, как правило. экспрессия генов при робертсоновских транслокациях не изменяются, поэтому виды практически неотличимы внешне. Однако они имеют разные кариотипы, а плодовитость при межвидовых скрещиваниях резко понижена.
ВСЕ ХРОМОСОМНЫЕ ПЕРЕСТРОЙКИ МОГУТ БЫТЬ ПОДРАЗДЕЛЕНЫ НА:
При сбалансированных перестройках изменяется порядок сегментов (локусов, генов) на хромосоме, но не происходит количественных нарушений генетического материала. (Например, инверсия и взаимные транслокации).
При возникновении несбалансированных перестроек всегда имеет место нарушение «дозы» определенных сегментов хромосомы. Это сопровождается изменением баланса генов и манифестацией той или иной формы хромосомной болезни.
СИНДРОМ ВОЛЬФА-ХИРШХОРНА (делеция короткого плеча хромосомы 4)
• Популяционная частота - 1:100000.
• обусловлен делецией сегмента короткого плеча хромосомы 4.
• Клинически характеризуется мВПР (микроцефалия, клювовидный нос, гипертелоризм, эпикант, аномальные ушные раковины, расщелины верхней губы и нёба, аномалии глазных яблок, антимонголоидный разрез глаз, маленький рот, пороки внутренних органов) с последующей резкой задержкой физического и психомоторного развития.
• Жизнеспособность детей резко снижена. Большинство умирают в возрасте до 1 года.
СИНДРОМ КОШАЧЬЕГО КРИКА (делеция короткого плеча хромосомы 5)
• Популяционная частот 1:45000 – 1:50000.
• Синдром обусловлен делецией короткого плеча 5 хромосомы.
• Кариотип: 46,ХХ, del 5p или 46,XY,del 5p.
• Специфический плач, напоминающий кошачье мяуканье или крик. Он обусловлен изменением гортани (сужение, мягкость хрящей, уменьшение надгортанника, необычная складчатость слизистой оболочки). С возрастом этот симптом исчезает.
СИНДРОМ ЧАСТИЧНОЙ ТРИСОМИИ ПО КОРОТКОМУ ПЛЕЧУ ХРОМОСОМЫ 9
• Клиническая картина многообразна и включает внутриутробные и постнатальные нарушения развития: задержку роста, умственную отсталость, микробрахицефалию, антимонголоидный разрез глаз, энофтальм (глубоко посаженные глаза), гипертелоризм, округлый кончик носа, опущенные углы рта, низко расположенные оттопыренные ушные раковины с уплощенным рисунком, гипоплазию (иногда дисплазию) ногтей. Врожденные пороки сердца обнаружены у 25% больных.
СИНДРОМ ПРАДЕРА-ВИЛЛИ И АНГЕЛЬМАНА
• у 70% больных наблюдается частичная делеция длинного плеча 15-й хромосомы (отцовская аллель), у 5% заболевание связано с другими перестройками хромосомы 15.
• Характерные внешние признаки: череп со сдавленной с боков лобной частью, миндалевидный разрез глаз, опущенные углы рта, маленькие стопы и кисти)
Наблюдается отставание умственного развития, поведенческие нарушения, задержка физического развития, низкорослость, гипотония, гипогонадизм.
Синдром Ангельмана – генетическое заболевание, для которого характерны психическая задержка развития, судорожные припадки, нарушения сна, хаотичные движении рук, частый смех и практически постоянная улыбка.
• Встречаемость 1:20 000
• Кариотип больных с синдромом Ангельмана — 46 XX или XY, 15р−.
• Продолжительность жизни 20-50 лет
1. Курчанов, Н. А. Генетика человека с основами общей генетики / Н.А. Курчанов. - М.: СпецЛит, 2009.
2. Гнатик, Е. Н. Генетика человека. Былое и грядущее / Е.Н. Гнатик. - М.: ЛКИ, 2010
3. Баранов, В. С. Цитогенетика эмбрионального развития человека / В. С. Баранов, Т. В. Кузнецова. -Спб. : Изд-во Н-Л, 2007. - 640 с.
4. Клаг У., Каммингс М. Основы генетики — М.: Мир, 2007.
5. Биология. Книга 1. Под ред. акад. РАМН Ярыгина В. Н. — М.: Высшая школа, 2003.
6. Грин Н. и др., Биология — М.: Мир, 1990. Т. 1-3.
7. Жимулев И. Ф. Общая и молекулярная генетика. — Новосибирск: Изд-во НГУ, 2003.
8. Р.В. Тузова, Н.А. Ковалев. Молекулярно-генетические механизмы эволюции органического мира. Генетическая и клеточная инженерия. – Минск: Беларусь, 2010
Хромосомные перестройки
- Хромосомные перестройки (хромосомные мутации, или хромосомные аберрации) — тип мутаций, которые изменяют структуру хромосом. Классифицируют следующие виды хромосомных перестроек: делеции (утрата участка хромосомы), инверсии (изменение порядка генов участка хромосомы на обратный), дупликации (повторение участка хромосомы), транслокации (перенос участка хромосомы на другую), а также дицентрические и кольцевые хромосомы. Известны также изохромосомы, несущие два одинаковых плеча. Если перестройка изменяет структуру одной хромосомы, то такую перестройку называют внутрихромосомной (инверсии, делеции, дупликации, кольцевые хромосомы), если же двух разных, то межхромосомной (дупликации, транслокации, дицентрические хромосомы). Хромосомные перестройки подразделяют также на сбалансированные и несбалансированные. Сбалансированные перестройки (инверсии, реципрокные транслокации) не приводят к потере или добавлению генетического материала при формировании, поэтому их носители, как правило, фенотипически нормальны. Несбалансированные перестройки (делеции и дупликации) меняют дозовое соотношение генов, и, как правило, их носительство сопряжено с существенными отклонениями от нормы.
Связанные понятия
Транслока́ция — тип хромосомных мутаций, при которых происходит перенос участка хромосомы на негомологичную хромосому. Отдельно выделяют реципрокные транслокации, при которых происходит взаимный обмен участками между хромосомами, и Робертсоновские транслокации, или центрические слияния, при которых происходит слияние акроцентрических хромосом с полной или частичной утратой материала коротких плеч.
Делеции (от лат. deletio — уничтожение) — хромосомные перестройки, при которых происходит потеря участка хромосомы. Делеция может быть следствием разрыва хромосомы или результатом неравного кроссинговера. По положению утерянного участка хромосомы делеции классифицируют на внутренние (интерстициальные) и концевые (терминальные).
Инве́рсия — хромосомная перестройка, при которой происходит поворот участка хромосомы на 180°. Инверсии являются сбалансированными внутрихромосомными перестройками. Различают парацентрические (инвертированный фрагмент лежит по одну сторону от центромеры) и перицентрические (центромера находится внутри инвертированного фрагмента) инверсии. Инверсии играют роль в эволюционном процессе, видообразовании и в нарушениях фертильности.
Дупликация (лат. duplicatio — удвоение) — разновидность хромосомных перестроек, при которой участок хромосомы оказывается удвоенным. Может произойти в результате неравного кроссинговера, ошибки при гомологичной рекомбинации, ретротранспозиции.
Митоти́ческий кроссинго́вер — тип генетической рекомбинации, который может проходить в соматических клетках при митотических делениях как у организмов, обладающих полом, так и бесполых организмов (например, некоторых одноклеточных грибов, у которых не известен половой процесс). В случае бесполых организмов митотическая рекомбинация является единственным ключом к пониманию сцепления генов, так как у таких организмов это единственный способ генетической рекомбинации. Кроме того, митотическая рекомбинация.
Гомологи́чные хромосо́мы — пара хромосом приблизительно равной длины, с одинаковым положением центромеры и дающие одинаковую картину при окрашивании. Их гены в соответствующих (идентичных) локусах представляют собой аллельные гены — аллели, то есть кодируют одни и те же белки или РНК. При двуполом размножении одна гомологичная хромосома наследуется организмом от матери, а другая — от отца.
Рекомбинация — перераспределение генетического материала (ДНК или РНК) путём разрыва и соединения разных молекул, приводящее к появлению новых комбинаций генов или других нуклеотидных последовательностей. В широком смысле слова включает в себя не только рекомбинацию между молекулами ДНК, но и перекомбинацию (сортировку) генетического материала на уровне целых хромосом или ядер, а также обмен плазмидами между клетками.
Точечная мутация — тип мутации в ДНК или РНК, для которого характерна замена одного азотистого основания другим. Термин также применяется и в отношении парных замен, инсерции или делеции одного или нескольких нуклеотидов. Точечные мутации, возникающие в некодирующей ДНК, обычно никак себя не проявляют. Точечный мутант — организм, в генотипе которого произошла точечная мутация.
Сцепленное наследование — феномен скоррелированного наследования определённых состояний генов, расположенных в одной хромосоме.
Кроссинго́вер (от англ. crossing over — пересечение) — процесс обмена участками гомологичных хромосом во время конъюгации в профазе первого деления мейоза, которое происходит, например, при образовании гамет или спор. Помимо мейотического, описан также митотический кроссинговер.
Псевдоаутосо́мные о́бласти (англ. pseudoautosomal region — PAR) — гомологичные участки половых хромосом различного типа; у млекопитающих они, соответственно, находятся на X-хромосоме и Y-хромосоме. Все гены, расположенные в этих областях, есть у обоих полов и наследуются так же, как и любые аутосомные гены, отсюда и название областей.
Мозаицизм (генетический мозаицизм, хромосомный мозаицизм — mosaicism; мозаичность; могут употребляться синонимы «мозаичная форма», «мозаичный кариотип») — от фр. mosaique «мозаика» — наличие в тканях (растения, животного, человека) генетически различающихся клеток.
Геномная нестабильность (англ. genomic instability) (также «генетической нестабильность» (англ. genetic instability) или «нестабильность генома» (англ. Genome instability)) определяется высокой частотой мутаций в геноме клеточной линии. Эти мутации могут включать в себя изменения в последовательности нуклеиновых кислот, хромосомные перестройки или анеуплоидию. Геномная нестабильность является центральным фактором канцерогенеза, но также фактором некоторых нейродегенеративных заболеваний, таких как.
Геномный импринтинг — эпигенетический процесс, при котором экспрессия определённых генов осуществляется в зависимости от того, от какого родителя поступили аллели. Наследование признаков, определяемых импринтируемыми генами, происходит не по Менделю. Импринтинг осуществляется посредством метилирования ДНК в промоторах, в результате чего транскрипция гена блокируется. Обычно импринтируемые гены образуют кластеры в геноме. Импринтинг некоторых генов в составе генома показан для насекомых, млекопитающих.
Локус (лат. locus — место) в генетике означает местоположение определённого гена на генетической или цитологической карте хромосомы. Вариант последовательности ДНК в данном локусе называется аллелью. Упорядоченный перечень локусов для какого-либо генома называется генетической картой.
Метилирование ДНК — это модификация молекулы ДНК без изменения самой нуклеотидной последовательности ДНК, что можно рассматривать как часть эпигенетической составляющей генома.
Теломе́ры (от др.-греч. τέλος — конец и μέρος — часть) — концевые участки хромосом. Теломерные участки хромосом характеризуются отсутствием способности к соединению с другими хромосомами или их фрагментами и выполняют защитную функцию.
Гомеобокс (англ. homeobox) — последовательность ДНК, обнаруженная в генах, вовлечённых в регуляцию развития у животных, грибов и растений. Эти гены кодируют факторы транскрипции, которые, как правило, переключают каскады других генов. Гомеобокс состоит приблизительно из 180 пар нуклеотидов и кодирует белковый домен длиной в 60 аминокислот (гомеодомен), который может связывать ДНК.
Ге́терозиго́тными называют диплоидные или полиплоидные ядра, клетки или многоклеточные организмы, копии генов, которые в гомологичных хромосомах представлены разными аллелями. Когда говорят, что данный организм гетерозиготен (или гетерозиготен по гену X), это означает, что копии генов (или данного гена) в каждой из гомологичных хромосом несколько отличаются друг от друга.
Алле́ли (от греч. ἀλλήλων — друг друга, взаимно) — различные формы одного и того же гена, расположенные в одинаковых участках (локусах) гомологичных хромосом. В диплоидном организме может быть два одинаковых аллеля одного гена, в этом случае организм называется гомозиготным, или два разных, что приводит к гетерозиготному организму. Термин «аллель» предложен В. Иогансеном (1909 г.).
Хромосомная нестабильность (англ. Chromosomal instability) (CIN) — один из видов геномной нестабильности, при которой в дочерних поколениях делящихся клеток наблюдаются неклональные изменения кариотипа, а именно: потери или приобретения хромосом и их участков.
Соматические клетки (др.-греч. σῶμα — тело) — клетки, составляющие тело (сому) многоклеточных организмов и не принимающие участия в половом размножении. Таким образом, это все клетки, кроме гамет.
Дозовая компенсация генов — эпигенетические механизмы, позволяющие уравнять уровень экспрессии сцепленных с полом генов у самцов и самок тех видов, в которых определение пола происходит с помощью половых хромосом. Так, например, у самцов млекопитающих гены X-хромосомы, не считая псевдоавтосомных областей, присутствуют в одной копии, а у самок — в двух. Поскольку такая разница могла бы привести к серьезным аномалиям, существуют механизмы дозовой компенсации генов, не связанных непосредственно с определением.
Хромосо́мные террито́рии — отдельные и почти не перекрывающиеся области, которые занимают хромосомы в ядре во время интерфазы клеточного цикла. Хромосомные территории имеют сферическую форму с диаметром от одного до нескольких микрометров. Считается, что территориальная организация хромосом характерна для всех эукариот, хотя есть и исключения, например, дрожжи Saccharomyces cerevisiae.
Дикий тип — наиболее часто встречающийся в природной популяции фенотип (или совокупность фенотипов); в селекции микроорганизмов — штамм, выделенный непосредственно из природного субстрата. Исходно этот термин использовался для обозначения продукта «нормального» аллеля, в противовес нестандартному продукту «мутантного» аллеля. Тем не менее, все «мутантные» аллели во всём своём разнообразии присутствуют в дикой популяции, и даже могут закрепиться как новый дикий тип в результате генетического дрейфа.
Транспозоны (англ. transposable element, transposon) — это участки ДНК организмов, способные к передвижению (транспозиции) и размножению в пределах генома. Транспозоны также известны под названием «прыгающие гены» и являются примерами мобильных генетических элементов.
Центромера — участок хромосомы, который связывает сестринские хроматиды, играет важную роль в процессе деления клеточного ядра и участвует в контроле экспрессии генов. Характеризуется специфическими последовательностью нуклеотидов и структурой.
Гомозиго́та (др.-греч. ὅμοιος «подобный, похожий, равный» + ζυγωτός «спаренный, удвоенный») — диплоидный организм или клетка, несущий идентичные аллели (аллельные гены) в гомологичных хромосомах (аа, или АА)
Эписта́з — тип взаимодействия генов, при котором проявление одного гена находится под влиянием другого гена (генов), неаллельного ему. Ген, подавляющий фенотипические проявления другого, называется эпистатическим (ингибитором, супрессором); ген, чья активность изменена или подавлена, называется гипостатическим.
Псевдогены (англ. pseudogenes) — нефункциональные аналоги структурных генов, утратившие способность кодировать белок и не экспрессирующиеся в клетке. Термин «псевдоген» был впервые предложен в 1977 году. Некоторые псевдогены могут копироваться из мРНК и включаться в хромосомы, такие последовательности называются процессированными псевдогенами (ретропсевдогенами). Тем не менее, они также нефункциональны. Псевдогены происходят от обычных функциональных генов, однако утрачивают способность экспрессии.
Нокаут гена (англ. gene knockout) — это метод молекулярной генетики, при котором из организма удаляют или делают неработоспособными определенные гены. Таким образом получают организм, «нокаутный» по неработающим генам. Нокаутные организмы помогают узнать функции генов, нуклеотидная последовательность которых известна (обратная генетика). Различия между нокаутным и нормальным организмом могут свидетельствовать о функции выключенного гена.Метод введения генов knock-in имеет сходства с методом gene.
Эпигенетическим наследованием называют наследуемые изменения в фенотипе или экспрессии генов, вызываемые механизмами, отличными от изменения последовательности ДНК (приставка эпи- означает в дополнение). Такие изменения могут оставаться видимыми в течение нескольких клеточных поколений или даже нескольких поколений живых существ.
Гены «домашнего хозяйства» (англ. housekeeping genes) — это гены, необходимые для поддержания важнейших жизненных функций организма, которые экспрессируются практически во всех тканях и клетках на относительно постоянном уровне. Гены домашнего хозяйства функционируют повсеместно, на всех стадиях жизненного цикла организма.
Си́напсис — конъюгация хромосом, попарное временное сближение гомологичных хромосом, во время которого между ними может произойти обмен гомологичными участками. Происходит в профазе I мейоза . Когда гомологичные хромосомы синаптируют, их концы прикрепляются к ядерной оболочке. Потом такие концевые мембранные комплексы перемещаются при содействии ядерного цитоскелета, пока соответственные концы хромосом не объединятся в пары. После этого и межконцевые участки хромосом начинают сближаться, при этом.
Мобильные генетические элементы (МГЭ, англ. Mobile genetic elements, MGE) — последовательности ДНК, которые могут перемещаться внутри генома.
Эпигенетика (др.-греч. ἐπι- — приставка, обозначающая пребывание на чём-либо или помещение на что-либо) — в биологии, в частности в генетике — представляет собой изучение закономерностей эпигенетического наследования — изменения экспрессии генов или фенотипа клетки, вызванных механизмами, не затрагивающими последовательности ДНК. Эпигенетические изменения сохраняются в ряде митотических делений соматических клеток, а также могут передаваться следующим поколениям. Примерами эпигенетических изменений.
Повреждение ДНК — это изменение химической структуры ДНК, такое как однонитевой или двунитевой разрыв сахаро-фосфатного остова ДНК, потеря или химическое изменение азотистых оснований, сшивка цепей ДНК, сшивка ДНК-белок. Структура ДНК в клетке регулярно нарушается из-за того, что при естественном метаболизме образуются соединения, которые обладают способностью повреждать ДНК. Эти соединения включают активные формы кислорода, реактивные формы азота, активные карбонильные группы, продукты перекисного.
Конденсины — большие белковые комплексы, которые играют главную роль в расхождении хромосом во время митоза и мейоза.
Синтения — по изначальному определению, расположение каких-либо локусов на одной и той же хромосоме (независимо от того, наблюдается ли у них сцепленное наследование). Сегодня, однако, синтенией обычно называют ситуацию, когда расположение каких-либо локусов на одной и той же хромосоме наблюдается в разных наборах хромосом (например, у разных видов). Это явление также называют общей синтенией (англ. shared synteny). Если при этом совпадает и порядок этих локусов в хромосоме, это называется коллинеарностью.
Мутационная теория канцерогенеза — учение, согласно которому причиной возникновения злокачественных опухолей являются мутационные изменения генома клетки. В настоящее время эта теория является общепринятой. В подавляющем большинстве случаев злокачественные новообразования развиваются из одной опухолевой клетки, то есть имеют моноклональное происхождение. Согласно современным представлениям, мутации, которые в конце концов приводят к развитию опухоли, могут иметь место как в половых (около 5 % всех.
Клетки зародышевой линии — клетки многоклеточного организма, дифференцированные или отделённые таким образом, что в обычных процессах воспроизведения они дают начало потомству.Как правило, такая передача осуществляется в процессе полового размножения; обычно это процесс, включающий систематические изменения генетического материала, изменения, которые возникают во время рекомбинации, мейоза и оплодотворение или сингамии например. Однако, существует много исключений, включая процессы, такие как различные.
Мута́ция (лат. mutatio — изменение) — стойкое (то есть такое, которое может быть унаследовано потомками данной клетки или организма) изменение генома. Термин предложен Гуго де Фризом в 1901 году. Процесс возникновения мутаций получил название мутагенеза.
Робертсоновская транслокация, или центрическое слияние, — хромосомная перестройка, при которой происходит слияние двух акроцентрических хромосом с образованием одной метацентрической или субметацентрической хромосомы. Слияние акроцентриков происходит в околоцентромерных районах, то есть в этой перестройке происходит транслокация целого плеча. Робертсоновские транслокации относятся к межхромосомным перестройкам. Робертсоновские транслокации играют роль в видообразовании, являются частым механизмом.
Сантиморган (сокращенно: сМ) в генетике — единица измерения генетических расстояний между полиморфными фрагментами генома (локусами или маркерами), которая определена как расстояние, на котором вероятность рекомбинации генов в мейозе составляет 1 %.
Гомологи́чная рекомбина́ция, или о́бщая рекомбина́ция, — тип генетической рекомбинации, во время которой происходит обмен нуклеотидными последовательностями между двумя похожими или идентичными хромосомами. Это наиболее широко используемый клетками способ устранения двух- или однонитевых повреждений ДНК. Гомологичная рекомбинация также создает разнообразие комбинаций генов во время мейоза, обеспечивающих высокий уровень наследственной изменчивости, что, в свою очередь, позволяет популяции лучше адаптироваться.
Наследование, сцепленное с полом — наследование какого-либо гена, находящегося в половых хромосомах. Наследование признаков, проявляющихся только у особей одного пола, но не определяемых генами, находящимися в половых хромосомах, называется наследованием, ограниченным полом.
Микрохромосо́мы — очень маленькие хромосомы, типичные для кариотипов птиц, некоторых рептилий, рыб и земноводных; у млекопитающих они, судя по всему, отсутствуют. Их размер составляет меньше 20 мегабаз; хромосомы, чей размер превышает 40 мегабаз, называются макрохромосомами, а хромосомы размером от 20 до 40 мегабаз — промежуточными хромосомами.
Генная конверсия представляет собой процесс, посредством которого одна последовательность ДНК заменяется гомологичной последовательностью, так что последовательности становятся идентичными. Конверсия гена может быть либо аллельной, что означает, что один аллель гена заменяет другой аллель того же гена, либо неаллельной/эктопической, при которой одна паралогичная ДНК-последовательность преобразует другую.Паралогичными называют такие последовательности, которые присутствуют в одном и том же геноме.
Флуоресце́нтная гибридиза́ция in situ, или метод FISH (англ. fluorescence in situ hybridization — FISH), — цитогенетический метод, который применяют для детекции и определения положения специфической последовательности ДНК на метафазных хромосомах или в интерфазных ядрах in situ. Кроме того, FISH используют для выявления специфических мРНК в образце ткани. В последнем случае метод FISH позволяет установить пространственно-временные особенности экспрессии генов в клетках и тканях.
Научная электронная библиотека

Хромосомные (геномные) мутации (аномалии) связаны либо с различными структурными перестройками хромосом, либо с изменением их числа (n). Численные изменения в наборе хромосом (кариотипе) могут быть двух типов: полиплоидии – умножение полного хромосомного набора (3n, 4n и т. д.) или генома, кратное гаплоидному числу хромосом; анеуплоидии – увеличение или уменьшение числа хромосом в наборе, некратное гаплоидному. Эти количественные изменения кариотипа обусловлены, как правило, нарушениями мейоза или митоза. Численные хромосомные аномалии в виде анеуплоидии делятся на моносомию (потерю хромосомы или её части – частичная моносомия) и трисомию или полисомию (приобретение одной/нескольких хромосом или её части – частичная трисомия). Данные изменения кариотипа связаны с комплексом врождённых пороков развития и, как правило, с заболеваниями, сопровождающимися умственной отсталостью или тяжелыми психическими расстройствами. В настоящее время описаны случаи изменений хромосомного набора с участием половых хромосом и некоторых аутосом при шизофрении и аутизме. Например, до 5–15 % детей с аутистическими расстройствами имеют хромосомные аномалии. Это позволяет рассматривать хромосомный дисбаланс клеток организма в качестве одной из возможных причин отдельных случаев нервных и психических болезней.
Структурные изменения могут затрагивать всю хромосому, а также сопровождаться изменением количества генетического материала в ядре или его перемещением. Сбалансированные хромосомные аномалии представляют собой перестройки, за счет которых выявляется кариотип с измененным набором расположения генов в пределах хромосом или между хромосомами, который отличается от нормального кариотипа. В большинстве случаев носители сбалансированных хромосомных аномалий фенотипически нормальны, но для их потомства возникает большой риск иметь несбалансированный кариотип. Следует отметить, что в отдельных случаях носители сбалансированного кариотипа могут иметь различные врождённые пороки и/или микроаномалии, а также нарушения нервного и психического развития. Если при структурных хромосомных мутациях наблюдается потеря или приобретение генетического материала, то они являются несбалансированными хромосомными аномалиями.
Цитогенетически структурные хромосомные перестройки классифицируют по принципу линейной последовательности расположения генов: делеции (потеря хромосомных участков), дупликации (удвоение хромосомных участков), инверсии (перевертывание на 180° относительно нормальной последовательности хромосомных участков), инсерции (вставки хромосомных участков) и транслокации (изменение расположения хромосомных участков). В последнее время в литературе хромосомные микроаномалии и перестройки могут обозначаться, как геномные.
Изменения генома (хромосом), приводящие к редким заболеваниям, могут включать как крупные микроскопически видимые перестройки (более 5 млн пн), так и вариации числа копий последовательностей ДНК (CNV) и однонуклеотидные полиморфные изменения последовательности ДНК (SNP). Как уже было сказано выше, в настоящее время для определения причины заболевания на геномном уровне используются различные технологии, наиболее распространенными из которых являются полногеномные методы, в частности, молекулярное кариотипирование (arrayCGH). Однако степень патогенности выявленных вариаций генома можно установить только при помощи использования инновационных биоинформатических технологий. Большой массив информации, собранный на интернет-ресурсах, позволяет уточнить функциональные особенности (онтологию) как отдельного гена, так и целой генной сети за счёт анализа последовательности кодируемого белка и моделирования молекулярных процессов, инициированных геномным изменением.
Таким образом, с хромосомными болезнями связаны аномалии микроскопически видимых численных или структурных нарушений хромосом, геномные же болезни связаны как с микроаномалиям хромосом, так и с вариациями числа копий последовательностей ДНК (CNV). С внедрением в клиническую практику современных высокоразрешающих молекулярных методов исследования генома появилась возможность выявлять вариации генома размером от 100 пн, мозаицизм низкого уровня и участки потери гетерозиготности. Размеры перестроек могут варьировать от тысячи до нескольких миллионов пн. При «крупных» перестройках в генонасыщенных участках может быть охвачено от нескольких десятков до нескольких сотен генов. В случае если перестройка небольшого размера и затрагивает область внутри одного или нескольких генов, её называют интрагенной. Как и в случае других типов генетических мутаций, некоторые CNV передаются по наследству, но большинство патогенных аномалий возникают de novo. Как было отмечено выше, структурные вариации генома можно разделить на рекуррентные (часто встречающиеся в популяции, предположительно непатогенные) и нерекуррентные (редко встречающиеся перестройки). Хотя последствия большинства CNV остаются неизвестными, появляется всё больше доказательств того, что генетические нарушения при наиболее распространённых психических и неврологических нарушениях включают в себя различные типы как часто встречающихся, так и редких генетических вариаций. Отдельное внимание стоит уделить биоинформатическому анализу (см ниже), который используется для интерпретации данных, полученных при молекулярном кариотипировании. С использованием данного подхода к анализу полученных молекулярно-цитогенетических результатов можно проводить приоритизацию генов-кандидатов и процессов, лежащих в основе развития патологии.
Большое значение имеет изучение хромосомных мутаций под действием факторов внешней среды. Показано, что хромосомы человека отличаются высокой чувствительностью к действию радиации и химических веществ, которые принято называть мутагенными факторами (мутагенами). При анализе воздействия этих факторов следует различать нарушения в соматических и половых клетках. Первые затрагивают непосредственно жизнедеятельность исследуемого организма, тогда как вторые проявляются в последующих поколениях. Мутации хромосом в зародышевых клетках ведут к образованию аберрантных гамет, в результате которых возможна гибель зигот, эмбрионов на ранних стадиях внутриутробного развития, а также рождение детей с специфическими хромосомными аномалиями, которые проявляются в виде определенной клинической картины или определенного фенотипа. Мутации хромосом в соматических клетках ведут к образованию неспецифичных хромосомных аномалий в виде хромосомных или хроматидных пробелов, разрывов, обменов в кариотипе, не ведущих к определенному фенотипу, характерному для конкретного наследственного заболевания. Подобные мутации не наследуются. Следует отметить, что при изучении такого рода воздействия мутагенных факторов представляется возможным оценить качественно и количественно действие ионизирующей радиации, химических веществ, вирусов, но полученные данные не могут быть перенесены на половые клетки, где результатом действия являются специфические хромосомные аномалии, влияющие на фенотип.
Хромосомные аномалии могут проявляться в так называемых мозаичных формах, к которым приводит неправильное деление клеток на различных стадиях эмбрионального и постнатального развития. Это позволяет разделить хромосомные аномалии на мозаичные и регулярные (аномальный кариотип наблюдается во всех клетках организма). Хромосомный мозаицизм представляет собой наличие нескольких популяций клеток с различным друг от друга хромосомным набором. Как правило, при мозаичных формах хромосомных аномалий наблюдают отсутствие отдельных клинических признаков определенного хромосомного синдрома и более легкое течение заболевания, но некоторые симптомы практически всегда присутствуют. Мозаичные структурные хромосомные аномалии наблюдаются достаточно редко, поэтому, когда речь идет о мозаичных хромосомных аномалиях, имеются в виду, в основном, численные аномалии, мозаичные формы которых имеют достаточно высокую популяционную частоту. Следует также отметить феномен тканеспецифического хромосомного мозаицизма, когда клетки с аномальным хромосомным набором присутствуют только в определенной ткани организма.
Читайте также: