Стадия морулы. Бластуляция зиготы
Обновлено: 18.04.2024
ООО Медицинский центр "МирА", Москва, Россия, 119334
Научный центр акушерства, гинекологии и перинатологии им. акад. В.И. Кулакова Минздрава России, Москва
ФГБУ «Научный центр акушерства, гинекологии и перинатологии им. В.И. Кулакова» Минздрава России, Москва
ФГБОУ ВО «Приволжский исследовательский медицинский университет» Министерства здравоохранения Российской Федерации, Нижний Новгород, Россия
Роль морфологической оценки ооцита и эмбриона при использовании ВРТ (обзор литературы)
Журнал: Проблемы репродукции. 2015;21(1): 54‑58
ООО Медицинский центр "МирА", Москва, Россия, 119334
В обзоре приведены данные о существующей классической практике визуальной морфологической оценки ооцитов и эмбрионов в современной клинической репродуктологии. Приведены структурно-функциональные временно-обусловленные особенности ооцита и эмбриона в условиях наблюдения in vitro, влияющие на наступление беременности в программе ЭКО.
ООО Медицинский центр "МирА", Москва, Россия, 119334
Научный центр акушерства, гинекологии и перинатологии им. акад. В.И. Кулакова Минздрава России, Москва
ФГБУ «Научный центр акушерства, гинекологии и перинатологии им. В.И. Кулакова» Минздрава России, Москва
ФГБОУ ВО «Приволжский исследовательский медицинский университет» Министерства здравоохранения Российской Федерации, Нижний Новгород, Россия
В настоящее время существуют различные системы оценки качества гамет и эмбрионов в условиях in vitro. При этом основная роль принадлежит морфологической оценке на эмбриологическом этапе программы экстракорпорального оплодотворения и переноса эмбрионов (ЭКО и ПЭ) [1].
Процессы раннего эмбрионального развития, по данным литературы, рассматриваются с позиции перехода от ооцита к эмбриону (oocyte-to-embryo transition), что обусловливает необходимость качественного отбора ооцитов на преконцепционном этапе с целью получения эмбрионов, имеющих высокий шанс к имплантации [2].
Цель настоящего обзора — продемонстрировать структурно-функциональные последовательно изменяющиеся морфологические особенности ооцитов, эмбрионов, полученных в результате оплодотворения, и обосновать необходимость проведения динамического наблюдения в программе ЭКО.
Важным аспектом программы ЭКО является получение зрелых ооцитов, находящихся на стадии мейоза II (МII), способных к оплодотворению in vitro [3, 4]. Существуют различные классификации, описывающие структурные особенности женских гамет. По данным отечественных источников [5], существует ограниченное количество работ, посвященных данной теме, при этом отсутствуют указания на взаимосвязь тех или иных критериев качества с основными показателями результативности ЭКО. Относительно недавно в клиническую практику стала внедряться система оценки ооцитов на стадии MII — MOMS (MII oocyte morphological score), которая продемонстрировала значимую взаимосвязь уровня баллов по шкале MOMS и наступления беременности [6].
Наиболее точную оценку ооцита возможно произвести после денудации ооцита из ооцит-кумулюсного комплекса в программе интрацитоплазматической инъекции сперматозоида в ооцит (ИКСИ) [6—8].
В циклах стимуляции суперовуляции встречаются ооциты с различными повреждениями [9]. Аномалии ооцита можно условно разделить на экстрацитоплазматические и цитоплазматические. К экстрацитоплазматическим относятся нарушение формы и организации прозрачной оболочки ооцита, дебрис в перивителлиновом пространстве, аномалии первого полярного тельца (расположение, увеличение в размере, фрагментация, дегенерация), а также нарушение симметрии цитоплазматической мембраны ооцита. К цитоплазматическим аномалиям, определяемым на световом уровне, относят зернистость цитоплазмы, агрегаты гладкого эндоплазматического ретикулума, рефрактерные тела, вакуоли, нарушение вязкости цитоплазмы [6—9]. При помощи поляризационной микроскопии возможна визуализация веретена деления [10]. Для качественного ооцита веретено деления ооцита стадии МII не должно быть в позиции перед первым полярным тельцем, при этом изменение температуры в пределах 1 °C может значительно изменить структуру веретена деления. У некоторых ооцитов, имеющих первое полярное тельце, может не быть видимого в поляризационном микроскопе веретена деления. Поляризационная микроскопия дает возможность оценки веретена деления и позволяет прогнозировать течение процесса оплодотворения и последующего дробления развившегося в результате оплодотворения эмбриона [11, 12].
Качество ооцита является важным прогностическим фактором, так как ядерная и цитоплазматическая зрелость клетки напрямую связана с эффективностью цикла ИКСИ [8, 13]. Основными причинами выявляемых изменений являются анеуплоидии ооцитов и гипоксия. Предполагают, что интрафолликулярная гипоксия ведет к снижению рН в течение мейоза, что отражается на формировании веретена деления, приводит к повышению частоты анеуплоидий [14].
В ранних исследованиях плоидности зрелых ооцитов стадии МII показано, что частота анеуплоидий составляет более 50% [10]. Анеуплоидии в ооците происходят вследствие нерасхождения хромосом, задержки анафазы, преждевременного движения центриолей в течение преовуляторного мейоза [15]. Вследствие хромосомных дефектов происходят специфические нарушения на клеточном уровне в виде цитоплазматического дисморфизма, описанного для ооцитов [10]. Значительное кластирование органелл коррелирует с высокой степенью анеуплоидии, другие аномалии связаны с нарушением сегрегации эндоплазматического ретикулума и митохондрий [16—18].
В качестве основной причины самопроизвольного прерывания беременности ранних сроков отмечают нарушение морфологии ооцита [10, 19]. При повторных неудачах программ ЭКО и ПЭ явление клеточного дисморфизма — аномалий устройства цитоплазмы, может быть причиной идиопатического бесплодия, в основе которого лежит хроническая анеуплоидия [20]. Присутствие в ооците генетических дефектов на хромосомном и геномном уровне в некоторых случаях может не оказывать влияние на фенотип эмбриона, который по морфологическим характеристикам может не уступать остальным нормальным эмбрионам на 2—3-и сутки дробления [14]. Сочетание световой и поляризационной микроскопии позволяет оценивать специфические морфологические характеристики полученных ооцитов [5, 8].
По данным последнего систематического обзора L. Rienzi и соавт. [6], анализ 50 публикаций за последние 15 лет продемонстрировал необходимость пристального внимания эмбриолога к комплексу наиболее значимых критериев качества ооцита — оценка ооцит-кумулюсного комплекса, прозрачной оболочки, перивителлинового пространства, морфологии первого полярного тельца, формы ооцита, структуры цитоплазмы, веретена деления. Рассматриваемые по отдельности, эти параметры имеют низкую диагностическую эффективность.
Оценка эмбриона по морфологическим параметрам
В настоящее время в клинической практике проводится оценка качества эмбрионов по морфологическим характеристикам на основании системы динамического наблюдения (sequential embryo selection, SES) [22, 23, 26—28].
Представляет интерес морфологическая оценка качества эмбрионов на основании кумулятивного эмбрионального счета (cumulative embryo score, CES), разработанного C. Steer и соавт. в 1992 г. и модифицированного в 2007 г. группой ученых под руководством J. Holte путем добавления к CES интегративного морфологического описания дробящихся эмбрионов [23, 27]. Данная классификация оценивает совокупность скорости дробления эмбрионов, симметричности бластомеров, степени цитоплазматической фрагментации, мультинуклеарности бластомеров и присваивает эмбриону баллы по каждому критерию. По данным L. Rienzi и соавт. [29], система MOMS имеет преимущества перед CES по частоте наступления беременности и родов.
Для оценки эмбриона 1-х суток дробления используются методики определения структуры «halo», морфологии пронуклеусов, предшественников ядрышек (nucleolar precursor bodies, NPB) [30, 31].
Исследования внутрицитоплазматического движения показали появление полупрозрачной зоны у перикортикальной мембраны («halo» — сияние). Этот динамический процесс, описанный как волнообразная активность, или цитоплазматическое волнение, происходящее в течение перисингамической стадии, свидетельствует о полноценности эмбриона на стадии зиготы [24].
Мужской и женский пронуклеусы появляются по периферии ооцита и вращаются относительно центра 16—18 ч после оплодотворения. Пронуклеусы должны иметь одинаковый размер. При различии в размерах пронуклеусов высока вероятность анеуплоидии [31]. Для NPB предложена методика оценки следующих параметров: числа, позиции, расположения, размера. Равное расположение относительно оси перекрытия пронуклеусов, оптимальное число (5—7), размер NPB коррелируют с возможностью развития эмбриона до стадии бластоцисты [15, 30, 32].
В течение периода развития ооцита, со стадии примордиального до MII, нуклеоли активно синтезируют протеины и распределяют их по определенным зонам ядра до включения собственного генома эмбриона. Нуклеоли являются центрами продукции мРНК [4]. Ростовые факторы и регуляторные белки также продуцируются в нуклеолях. Эти участки ядра являются нуклеолярными организационными регионами (nucleolar organization regions, NOR) ядра. Гетерохроматин пяти хромосом 13, 14, 15, 21, 22 сцеплен с NOR. Именно с этими хромосомами связана наибольшая частота анеуплоидий, что подчеркивает важность строгой компартментализации ядра, определяющей качество ооцита и эмбриона [32, 33]. Нуклеоли состоят из трех функциональных компонент:
— плотного фибриллярного комплекса DFC (dense fibrillic complex), который важен для процесса транскрипции;
— фибриллярного комплекса FC (fibrillic component), который участвует в инактивации транскрипционных факторов;
— гранулярно-цитоплазматического комплекса GC (granular-cytoplasmatic component) [4].
Визуализация данных структур свидетельствует о функционировании NPB (так как они и являются собственно FC регионами) и указывает на присутствие гетерохроматина хромосом 13, 14, 15, 21, 22 в веретене деления. NPB появляются, когда хроматин конденсируется в митотическое веретено, что происходит к моменту пенетрации сперматозоида. Появление одного пронуклеуса может означать быструю или, наоборот, отложенную конденсацию хроматина в ядре, что связано с аномалиями развития, а именно анеуплоидиями [5, 10, 25, 29, 31, 34].
Работами многих авторов [29, 31] показано, что именно 1-е сутки дробления определяют качество эмбриона. Оптимальным является время вступления в первое митотическое деление через 23—24 ч после инсеминации ооцитов, однако оно может варьировать в сторону увеличения до 4 ч, что не является плохим прогностическим признаком [29]. После проникновения сперматозоида формируются пронуклеусы, оплодотворение завершается первым митотическим делением и формированием двухклеточного эмбриона.
Расположение веретена деления влияет на дальнейшее дробление. Если веретено поляризовано, то это может приводить к формированию эмбриона с неравными по размеру бластомерами. Аномальный цитокинез может приводить к патологической бластуляции, выражающейся в различиях размеров клеток во внутренней клеточной массе (inner cell mass, ICM) более чем на 30%, дефектам дальнейшего развития [10, 22, 35, 36]. На четырехклеточной стадии один из четырех бластомеров продолжает свое развитие в клетки, продуцирующие хорионический гонадотропин (ХГ), что является сигналом для эндометрия и подготовки к имплантации [10]. Достижение эмбрионом стадии четырех бластомеров происходит через короткую трехклеточную стадию. Длительная задержка на стадии трех, четырех бластомеров является неблагоприятным прогностическим признаком [23]. Важным для развития эмбриона является наличие ядра в каждом бластомере. Отсутствие ядра может свидетельствовать о том, что бластомер находится в переходной стадии. Через 42—44 ч после оплодотворения все бластомеры должны иметь ядра. Фрагментация ядер бластомеров коррелирует с низким качеством эмбриона и неблагоприятным исходом ЭКО и ПЭ [22, 30, 31]. Увеличение количества ядер происходит при аномальном кариокинезе или вследствие аномальной сегрегации хромосом, более 70% мультинуклеарных эмбрионов являются анеуплоидными [30].
По достижении четырехклеточной стадии бластомеры уменьшаются в 2 раза и до стадии восьми бластомеров не увеличиваются в размерах. Оценка эмбриона происходит на 3-и сутки дробления (64—66 ч после инсеминации ооцитов) в течение перехода от шести к восьми бластомерам, что может происходить очень быстро. На этой стадии исследуют число бластомеров (в норме 6—8), фрагментацию (в норме отсутствует или менее 10%), размер и соотношение бластомеров, количество ядер в бластомерах [29, 37].
Для 4-х суток дробления (66—112 ч после инсеминации ооцитов) характерной чертой эмбрионов является компактизация бластомеров и формирование морулы. Работами многих авторов показано, что ранняя компактизация и достижение стадии морулы в конце 3-х суток дробления, а также раннее начало процесса кавитации (формирования полостей между клетками морулы) являются благоприятными прогностическими признаками в отношении хорошего качества эмбриона [38].
На 5-е сутки дробления (113—116 ч после инсеминации ооцитов) происходит формирование бластоцисты. Работами многих авторов показана связь морфологии бластоцисты с исходом программ ВРТ [30, 32]. Эмбрион, развивающийся аномально в 1—3-и сутки дробления, может формировать нормальную по морфологии бластоцисту [39]. Признаками бластоцисты хорошего качества являются неповрежденные клетки, отсутствие гранулярных включений, хороший контакт между клетками, четкие контуры у 80% клеток, оптимальное соотношение бластоцели и клеточной массы. Наиболее используемой в настоящее время является классификация бластоцист по D. Gardner и W. Schoolcraft [40].
Морфологические критерии для ооцитов и эмбрионов постоянно дополняются и совершенствуются. Несмотря на это, по мнению J. Gerris и соавт. [10], эмбрионы с анеуплоидиями in vitro имеют характеристики, сходные с эуплоидными.
Наряду с морфологической оценкой в клиническую практику внедряются новые технологии, направленные на улучшение селекции эмбриона. Среди них можно выделить инвазивные и неинвазивные методы. К инвазивным относится преимплантационная генетическая диагностика, осуществляемая с использованием методов флюоресцентной гибридизации in situ (FISH), полимеразной цепной реакции с обратной транскрипцией (ОТ-ПЦР), анализа микроокружения, сравнительной геномной гибридизации. Объектами исследования при применении данных методов являются: полярное тельце ооцита, бластомер или клетки трофэктодермы эмбриона, ядро сперматозоида. Данные методы доказали свою эффективность и нашли применение в практике у пациенток старшего репродуктивного возраста при неоднократных неэффективных попытках ЭКО. Вместе с тем продолжают развиваться неинвазивные методы, позволяющие выбрать для переноса эмбрионы, имеющие высокий имплантационный потенциал. В настоящее время в работу многих центров репродуктивной медицины начинают внедряться системы непрерывного автоматического наблюдения за эмбрионами, которые, оценивая параметры цитокинеза, митоза, позволяют на основе многофакторного анализа выбрать наиболее перспективный эмбрион с точки зрения дальнейшего развития. Применяются технологии «-омик»: геномика, транскриптомика, протеомика, метаболомика. По мнению E. Forman и соавт. [41], полный геномный скрининг при селективном переносе эмбриона улучшит результаты ВРТ и снизит частоту невынашивания беременности.
Таким образом, анализ данных литературы показал, что морфологическая оценка играет основную роль в определении качества ооцитов и эмбрионов в процессе динамического наблюдения in vitro. Несмотря на это, субъективная оценка должна дополняться данными, характеризующими гаметы и эмбрион с морфофункциональной позиции. Внедрение технологий непрерывного наблюдения за эмбрионами, в основе работы которых лежит система фиксации структурных изменений эмбрионов во времени, позволит автоматизировать процесс отбора эмбрионов и дополнит классическую морфологическую оценку. Применение современных методов позволяет комплексно подходить к выбору эмбриона с высоким потенциалом к имплантации в программе ЭКО и ПЭ, принимая во внимание исходный статус гамет и клинико-анамнестические данные супружеских пар с бесплодием.
Стадия морулы. Бластуляция зиготы
Стадия морулы. Бластуляция зиготы
Последующие четыре дробления, при которых все восемь бластомеров снова делятся одновременно, характеризуются опять таки меридиональными бороздами, в результате чего возникает стадия развития, состоящая из шестнадцати бластомеров. При пятом, шестом и седьмом дроблениях их количество снова повторно удваивается, таким образом последовательно возникают образования, состоящие из 32, 64 и 128 бластомеров.
Борозды при этих делениях проходят в плоскостях, расположенных между полюсами и экватором по типу параллелей на земном шаре, поэтому они называются латитудинальными бороздами (latitudo = широта; параллели определяют географические широты). Начиная с восьмого дробления, прекращается одновременное деление всех бластомеров, они начинают делиться по группам, в связи с чем ход борозд по поверхности клеточного узла, начиная с данной стадии, перестает быть равномерным.
В результате описанного процесса из первоначальной одной клетки (оплодотворенной яйцевой клетки) возникает шароподобное образование, по величине намного отличающееся от оплодотворенного яйца и состоящее из значительного количества (нескольких сотен) мелких закругленных клеток. Поверхностные клетки при этом выступают над поверхностью образования наподобие отдельных закругленных частей малинового или тутового плода.
Из-за такого своего вида возникший в результате дробления клеточный узел называется морулой.

Стадия морулы является переходной стадией в последовательных процессах развития зародыша. При непрерывно продолжающемся дроблении бластомеров зародыш в отдельных отделах морулы начинает постепенно увеличиваться. Одновременно щели между клетками, вначале маленькие, в центре морулы начинают сливаться, образуя более крупные полости. Некоторые клетки в центральном отделе клеточного узла погибают и распадаются; таким образом, в результате этих процессов возникает более обширная сплошная полость.
Ее объем постепенно увеличивается, в то время как поверхностные бластомеры зародыша распределяются наподобие эпителия в виде сплошной однослойной клеточной стенки, окружающей центральную полость. Первоначальные борозды, проходящие между поверхностными клетками, исчезают, поскольку бластомеры на поверхности становятся более плоскими и своими боковыми стенками тесно примыкают друг к другу, в результате чего их форма адаптируется по типу цилиндрического, или же кубического эпителия.
Таким образом, возникает стадия развития, которая имеет вид маленького пузырька, ограниченного однослойной клеточной стенкой (бластодермой), со сравнительно большой центральной полостью (бластоцелем), заполненной водянистой жидкостью. Эта стадия развития называется бластулой.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Эмбриональное развитие
От момента образования зиготы и до выхода зародыша из яйцевых оболочек длится эмбриональный период развития.

Дробление зиготы
После того, как произошло оплодотворение - слияние сперматозоида и яйцеклетки, образовавшаяся зигота начинает интенсивно делиться. Ее множественные митотические деления называют дроблением.
Важная особенность дробления в том, что не происходит увеличение в размере зародыша: клетки дробятся (делятся) настолько быстро, что не успевают накопить цитоплазматическую массу. Дробление зиготы человека является полным неравномерным асинхронным.

В результате дробления образуется морула. Морула (лат. morum - ягода тутового дерева) - клетка на стадии этапа дробления, когда зародыш представляет собой компактную совокупность клеток (без полости внутри).
Бластуляция
Бластуляция - заключительный период дробления, в который зародыш называется бластулой.
После очередных этапов многократного деления образуется однослойный зародыш с полостью внутри - бластула (греч. blastos — зачаток).
Стенки бластулы состоят из бластомеров, которые окружают центральную полость - бластоцель (греч. koilos — полый). Соединяясь друг с другом, бластомеры образуют бластодерму из одного слоя клеток.

Гаструляция (греч. gaster — желудок, чрево)
Гаструляцией называют стадию эмбрионального развития, в ходе которой клетки, возникшие в результате дробления зиготы, формируют три зародышевых листка: эктодерму, мезодерму и энтодерму.
Стенка бластулы начинает впячиваться внутрь - происходит инвагинация стенки. По итогу такого впячивания зародыш становится двухслойным. Двухслойный зародыш называется - гаструла. Полость гаструлы называется гастроцель (полость первичной кишки), а отверстие, соединяющее гастроцель и внешнюю среду - первичный рот (бластопор).

У первичноротых животных на месте первичного рта (бластопора) образуется ротовое отверстие. К первичноротым относятся: кишечнополостные, плоские, круглые и кольчатые черви, моллюски, членистоногие.
У вторичноротых на месте бластопора формируется анальное отверстие, а ротовое отверстие образуется на противоположном полюсе. К вторичноротым относят хордовых и иглокожих (морских звезд, морских ежей).

При впячивании части бластулы (инвагинации) клетки бластодермы мигрируют внутрь и становятся энтодермой (греч. entós — внутренний). Оставшаяся часть бластодермы снаружи называется эктодермой (греч. ἔκτος - наружный).
Между энто- и эктодермой из группы клеток формируется третий зародышевый листок - мезодерма (греч. μέσος — средний).
Нейрула
Эта стадия следует за гаструлой. Ранняя нейрула представляет собой трехслойный зародыш, состоящий из энто-, экто- и мезодермы. На этапе нейрулы происходит закладка отдельных органов.
Важно отметить, что на стадии нейрулы происходит процесс нейруляции - закладывание нервной трубки. Нервная пластинка, образовавшаяся на ранних этапах, прогибается внутрь, при этом ее края сближаются и, замыкаясь, формируют нервную трубку.

Итак, как уже было сказано, на стадии нейрулы закладываются отдельные органы. Эктодерма образует покровный эпителий и нервную пластинку, мезодерма (из которой в дальнейшем появятся все соединительные ткани), энтодерма - окружает полость первичной кишки (гастроцель), образуя кишечник. От энтодермы отшнуровывается хорда.

Все три зародышевых листка требуют нашего особого внимания, а также понимания того, какие органы и структуры из них образуются.
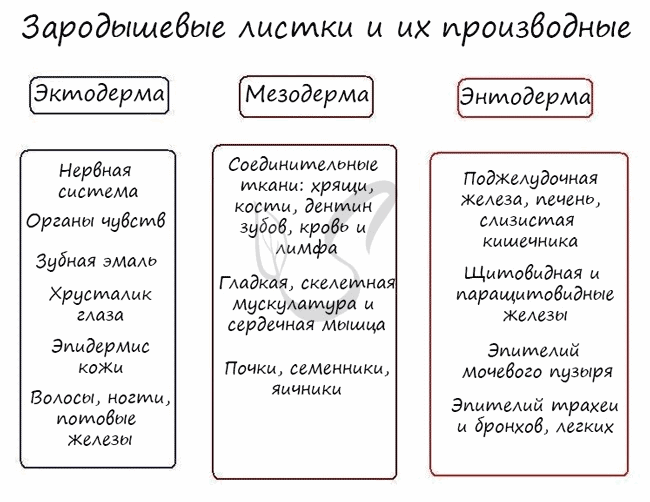
Эктодерма (греч. ἔκτος - наружный) - наружный зародышевый листок, образует головной и спинной мозг, органы чувств, периферические нервы, эпителий кожи, эмаль зубов, эпителий ротовой полости, эпителий промежуточного и анального отделов прямой кишки, гипофиз, гипоталамус.
Мезодерма (греч. μέσος — средний) - средний зародышевый листок, образует соединительные ткани: кровеносную и лимфатическую системы, костную и хрящевую ткань, мышечные ткани, дентин и цемент зубов, а также выделительную (почки) и половую системы (семенники, яичники).
Энтодерма (греч. entós — «внутренний») - внутренний зародышевый листок, образует эпителий пищевода, желудка, кишечника, трахеи, бронхов, легких, желчного пузыря, мочевого пузыря и мочеиспускательного канала, печень и поджелудочную железу, щитовидную и паращитовидную железы.
Из зародышевых листков образуются ткани, органы и системы органов. Такой процесс называется органогенезом. В период закладки органов важное значение имеет воздержание матери от вредных привычек (алкоголь, курение), которые могут нарушить процесс дифференцировки клеток и привести к тяжелейшим аномалиям, уродствам плода.
Некоторые лекарства также могут оказывать на плод тератогенный эффект (греч. τέρας — чудовище, урод), приводя к развитию уродств. Периоды закладки органов и систем органов вследствие их большой важности носят название критических периодов эмбриогенеза.

Анамнии и амниоты
Анамнии, или низшие позвоночные - группа животных, не имеющая зародышевых оболочек (зародышевого органа - аллантоиса и амниона). Анамнии проводят большую часть жизни в воде, без которой невозможно их размножение.
К анамниям относятся рыбы, земноводные.

Амниоты - группа высших позвоночных, характеризующаяся наличием зародышевых оболочек. К амниотам относятся пресмыкающиеся, птицы и млекопитающие.
Зародышевый орган, аллантоис, является органом дыхания и выделения.
За счет особых оболочек, развивающихся в ходе эмбрионального развития, амниона и серозы, у амниот формируется амниотическая полость. В ней находится зародыш, окруженный околоплодными водами. Благодаря такому гениальному устройству, амниотам для размножения и развития более не нужно постоянное нахождение в водоеме, они "обрели независимость" от него.

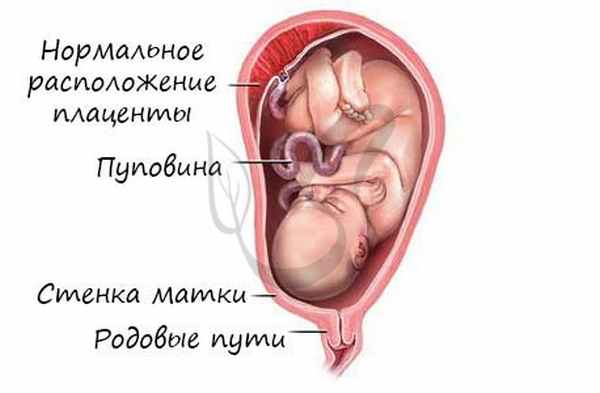
Развитие плода происходит в мышечном органе - матке, которая, сокращаясь во время родов, стимулирует изгнание плода через родовые пути. Питание осуществляется через плаценту - "детское место" - орган, который с одной стороны омывается кровью матери, а с другой - кровью плода. Через плаценту происходит транспорт питательных веществ и газообмен.
Соединяет плаценту и плод особый орган - пуповина, внутри которой проходят артерии, вены.
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Этапы и стадии эмбрионального периода развития
В биологии формирование индивидуального организма на протяжении всей жизни называется онтогенезом. На начальном этапе в него входят стадии эмбрионального периода развития (эмбриогенеза).
В биологии формирование индивидуального организма на протяжении всей жизни называется онтогенезом. На начальном этапе в него входят стадии эмбрионального периода развития (эмбриогенеза).

Этот процесс имеет важное значение в становлении животных и человека, так как в это время происходит закладка всех основ, объединение материнской и отцовской наследственности.
Общие сведения
В предэмбриональный период происходит формирование половых клеток, которые обладают гаплоидным набором хромосом и участвуют в размножении. Эмбриогенез наступает с оплодотворения женской яйцеклетки, когда берет начало зародышевый этап. В таблице кратко показаны основные ступени эмбрионального развития.
| Определение фазы | Особенности этапа |
| Образование зиготы | Результат соединения сперматозоида мужской особи с женской клеткой |
| Становление бластулы | Начало клеточного дробления. Образование многоклеточного шара с бластоцелью внутри |
| Стадия — гаструла | В результате деления клеток на одном из полюсов бластулы формируются зародышевые листы |
| Образование нейрулы | Становление у эмбриона нервной трубки и хорды |
| Органогенез | Начальный этап формирования органов |
После оплодотворения формируется зигота, какую можно описать как первый диплоидный элемент зародыша. В дальнейшем происходит дробление клетки, она перемещается по трубам материнского организма и фиксируется в маточной полости.

В результате дробления образуется многоклеточный эмбрион, который, благодаря дифференциации клеток, становится трехслойным зародышем. Стоит отметить, что при неблагоприятных условиях зигота впадает в спячку, а развитие может остановиться.
У человека, как и у всех млекопитающих, стадии эмбриогенеза охватывают период созревания зародыша внутри утробы матери до становления органов, частей тела, созревания плаценты.
На заключительных этапах развитие протекает при сформированных структурах, а зародыш становится плодом. Например, у крокодилов эмбриональное развитие заканчивается выходом особи из яйца.
Деление зиготы
На начальном этапе образования эмбриона происходит многократное дробление зиготы. Зародыш в этот период не меняется в размерах, так как образовавшиеся клетки во столько же раз меньше материнской, сколько сформировалось всего элементов. В результате сформировывается морула, которая представляет собой некий кластер с мелкими частичками зародыша.
Основные цели последовательных митотических делений:
- образование необходимого количества клеток для формирования органов;
- разделение цитоплазмы между первоначальными клетками;
- обеспечение нормального соотношения ядер и плазмы;
- определение осей организма.

Когда появится от 8 до 16 новых клеток, кластер разрастается за счет увеличения расстояния между частицами. Далее дистанция между ними сокращается, так как межклеточное пространство уменьшается. Существует два вида дробления:
- Голобластическое — происходит у млекопитающих, в том числе у людей, так как зародыш начинает питаться от организма матери.
- Меробластическое — развивается в замедленном темпе, характерно для птиц и пресмыкающихся.
При делении клеток образовываются прочные связи между элементами, которые затем используются для обмена сигналами.
В период изменения размеров кластера последовательное митотическое деление зиготы приостанавливается, а возобновляется оно после уплотнения зародыша. Вновь образованные клетки становятся предшественниками бластулы.
Формирование бластул и гаструляция
Как только прекращается деление зиготы, начинается фаза бластуляции. На этой ступени развития осуществляется передача информации с ДНК на РНК. Связанные клетки перестраиваются, формируется клеточная масса и трофобласт, который предохраняет зародыш от нежелательных воздействий.
Из клеточной массы образуется эмбрион, а при повторном делении клеток синтезируется ДНК. При этом используются ферменты, накопленные в период развития материнской половой клетки. В период становления бластулы зародыш не увеличивается, а происходит видоизменение зиготы на основе односменного зародыша.

Суть гаструляции состоит в образовании многослойного эмбриона, а этап зародышевого развития называется гаструлой.
Появление двух листков сигнализирует о начале гаструляции, а у позвоночных есть и третий слой, который называется мезодермой.
Эта стадия эмбрионального развития включает в себя несколько периодов, а именно:
- образование экто- и энтодермы в форме двухслойной бластулы;
- формирование мезодермы в виде трехслойного зародыша.
У позвоночных организмов эмбриогенез протекает быстрее, благодаря развитию среднего листка. У примитивных хордомезодерма появляется только на следующей стадии. При изменении периода развития у потомков, которая называется гетерохронией, возможны смещения начала образования главных органов. Существует несколько вариантов гаструляции:
- Инвагинация — область бластодермы расположена пластом внутрь. Встречается только в яйцеклетках с небольшим объемом желтка.
- Эпиболия — наличие на крупных клетках маленьких малоподвижных элементов, отстающих в скорости деления.
- Деноминация — представляет собой разделение бластодермы на разные слои. Происходит у живых организмов, которые откладывают яйца.
- Иммиграция — перемещение клеточных групп, расположенных в разных слоях. Бывает у всех видов млекопитающих.
При перестановке клеток наблюдается изменение строения эмбриона, который трансформируется из бластулы в гаструлу. Особое значение имеет дифференцировка, осуществляющая переход к реализации генетической программы.
Завершающий и критические этапы
Заключительной стадией развития эмбриогенеза является органогенез. В этот период развития зародыша происходят процессы нейруляции, гистогенеза и формирования органов. Сначала образуется нейрула, которая содержит три зародышевых слоя. Последний из них раздробляется на парные метамерные образования и осевые органы.
При гистогенезе формируются тканевые клетки тела. Так, наружный слой кожи и ткань нервной системы образуются из эктодермы, в результате чего появляется рефлекторная система, кожа и чувствительные рецепторы. Из энтодермы формируется эпителий, дающий начало становлению сосудов, легких и слизистых оболочек.

При зародышевом развитии происходит реакция эмбриона на отрицательное воздействие окружающей среды. Такие периоды носят название критических. Первый наступает сразу после оплодотворения и длится до 8 дней, то есть до фиксации клетки внутри матки. Ситуация эта опасна тем, что отсутствует связь с материнским организмом.
Если возникнут нежелательные условия, то зародыш может погибнуть. Следующий критический этап длится от 1 до 8 недель. При возникновении сложных ситуаций в период становления органов могут наблюдаться некоторые искажения в развитии.
Третий период наступает во время образования хориона и плаценты. Негативные явления на этой стадии могут нарушить формирование органов дыхания и повлиять на формирование плаценты. Следующий нежелательный этап протекает на 12—14 неделе развития.
В это время могут появиться некоторые дефекты в становлении половых органов у плода. Заключительная стадия приходится на 18—22 неделю, когда происходит завершение формирования нервной системы, рефлекторных реакций и вырабатываются некоторые виды гормонов.
После завершения зародышевого этапа наступает пренатальное становление плода, которое заканчивается рождением ребенка. С этого момента начинается постэмбриональный период, который длится до самой смерти.
Морула
Мо́рула (лат. morula — шелковица) — это стадия раннего эмбрионального развития зародыша, которая начинается с завершением дробления зиготы. Клетки морулы делятся гомобластически. После нескольких делений клетки зародыша формируют шаровидную структуру, напоминающий ягоду шелковицы.
В дальнейшем внутри зародыша появляется полость — бластоцель. Этот этап развития называется бластула.
См. также
Wikimedia Foundation . 2010 .
Полезное
Смотреть что такое "Морула" в других словарях:
МОРУЛА — МОРУЛА, ранняя стадия развития зародыша животных, на которой клетки находятся в процессе расщепления. Предшествует БЛАСТУЛЕ. Морула состоит из некоторого количества бластомеров, клеток, образовавшихся из оплодотворенной ЯЙЦЕКЛЕТКИ в результате… … Научно-технический энциклопедический словарь
МОРУЛА — (новолат. morula, от лат. morum тутовая ягода), стадия зародышевого развития нек рых губок, кишечнополостных, плоских червей, членистоногих, большинства млекопитающих в период дробления. Ранее М. считали стадией развития, предшествующей бластуле; … Биологический энциклопедический словарь
Морула — стадия эритроцитарного цикла развития плазмодия, в к рой взрослый шизонт приступает к делению на мерозоиты. (Источник: «Словарь терминов микробиологии») … Словарь микробиологии
морула — сущ., кол во синонимов: 1 • стадия (45) Словарь синонимов ASIS. В.Н. Тришин. 2013 … Словарь синонимов
МОРУЛА — (уменьш. от лат. morum ягода тутового дерева), эмбриологический термин, введенный Геккелем (Heckel) для обозначения заключительного стадия дробления, когда зародыш представляет собой компактную кучку клеток (бластомеров). В типичных случаях (яйца … Большая медицинская энциклопедия
морула — (лат. morula уменъш, от morum тутовая ягода) биол. стадия развития зародыша многоклеточных животных; представляет собой скопление клеток, напоминающее по внешнему виду ягоды тутового дерева или малины, без ясно обособившейся полости между ними;… … Словарь иностранных слов русского языка
морула — ЭМБРИОЛОГИЯ ЖИВОТНЫХ МОРУЛА – одна из стадий дробления некоторых беспозвоночных, членистоногих и большинства млекопитающих. На стадии морулы зародыш представляет собой похожий на тутовую ягоду шарик из плотно прилегающих друг к другу бластомеров … Общая эмбриология: Терминологический словарь
морула — morula морула. Pанняя стадия индивидуального развития, выделяемая в эмбриогенезе некоторых животных (иногда М. рассматривают, как один из типов бластулы ), представляет собой скопление бластомеров без внутренней полости. (Источник … Молекулярная биология и генетика. Толковый словарь.
МОРУЛА — (morula) ранняя стадия эмбрионального развития, возникающая в результате деления оплодотворенной яйцеклетки. На этой стадии эмбрион представляет собой скопление (наподобие ягоды тутового дерева ред.) клеток; морула является промежуточной стадией… … Толковый словарь по медицине
Морула (Morula) — ранняя стадия эмбрионального развития, возникающая в результате деления оплодотворенной яйцеклетки. На этой стадии эмбрион представляет собой скопление (наподобие ягоды тутового дерева ред.) клеток; морула является промежуточной стадией между… … Медицинские термины
Читайте также: