Цитоплазма бактерий. Бактериальный геном. Бактериальные рибосомы. Запасные гранулы бактерии.
Обновлено: 25.04.2024
Оглавление темы "Анатомия бактериальной клетки. Физиология бактерий.":
1. Анатомия бактериальной клетки. Поверхностные структуры бактерии. Капсула бактерий. Организация капсул. Окраска капсул бактерий. Состав капсул. Антигенные свойства капсул.
3. Микроворсинки бактерий. Фимбрии бактерий. F-пили (секс-пили) бактерии. Клеточная оболочка бактерий. Гликокаликс.
4. Клеточная стенка бактерий. Функции клеточной стенки. Строение клеточной стенки бактерии. Пептидогликан. Муреиновый мешок. Структура пептидогликана (муреина)
5. Грамотрицательные бактерии. Клеточная стенка грамотрицательных бактерий. Строение клеточной стенки грамотрицательных бактерий.
6. Грамположительные бактерии. Клеточная стенка грамположительных бактерий. Строение клеточной стенки грамположительных бактерий. Аутолизины бактерий. Сферопласты. Протопласты.
7. Цитоплазматическая мембрана (ЦПМ) бактерии. Состав цитоплазматической мембраны бактерий. Транспортные системы. Мезосомы. Периплазматическое пространство.
8. Цитоплазма бактерий. Бактериальный геном. Бактериальные рибосомы. Запасные гранулы бактерии.
9. Физиология бактерий. Питание бактерий. Тип питания бактерии. Голозои. Голофиты. Вода. Значимость воды для бактерий.
10. Усваиваемые бактериальной клеткой соединения. Пути поступления веществ в бактериальную клетку. Пассивный перенос. Диффузия.
По характеру движения подвижные бактерии разделяют на плавающие и скользящие (ползающие ). Орган движения плавающих бактерий - жгутики ; подвижность скользящих бактерий обеспечивают волнообразные сокращения тела.
Расположение жгутиков - характерный признак, имеющий таксономическое значение. Варианты расположения жгутиков приведены на рис. 4-1. У некоторых бактерий жгутики расположены по всей поверхности клеточной стенки (например, у бактерий рода Proteus), такие бактерии известны как перитрихи [от греч. peri-, вокруг, + trichos, волос]. Некоторые бактерии снабжены только одним толстым жгутиком (например, представители рода Vibrio), они известны как монотрихи . Политрихи - бактерии, имеющие одиночный по виду жгутик, образованный пучком из 2-50 жгутиков. Полярные жгутики прикреплены к одному или обоим концам бактерии. Монополярно-политрихиальное расположение жгутиков имеют лофотрихи [от греч. lophos, пучок, + trichos, волос], к ним, например, относят представителей рода Pseudomonas. Биполярно-политрихиальное жгутикование имеют амфитрихи [от греч. amphi-, двусторонний, + trichos, волос] (например, бактерии рода Spirillum).
Рис. 4-1. Варианты расположения жгутиков (вверху) и движений бактерий (внизу) .
Жгутик - спирально изогнутая полая нить, образованная субъединицами флагеллина. У разных бактерий толщина жгутиков варьирует от 12 до 18 нм, что составляет не более 1/10 диаметра жгутиков водорослей и простейших. Жгутики также различают по длине и диаметру витка. Место прикрепления жгутика к бактериальной клетке имеет сложное строение и состоит из базальной структуры и так называемого «крюка» (рис. 4-2). У грамположительных бактерий в состав базальной структуры входит одна пара, а у грамотрицательных бактерий - две пары колец. Кольца играют роль «приводного диска» и «подшипника». Вся конструкция выполняет функцию хемомеханического преобразователя (флагеллиновый мотор). У спирохет за движение ответственна особая органел-ла - осевая нить, состоящая из двух рядов бактериальных жгутиков, расположенных продольно внутри клетки.
Бактериальные жгутики совершают поступательные и вращательные движения, проталкивая бактерии через среду подобно корабельному винту. Они также могут изменять направление вращения и тянуть клетку подобно пропеллеру. Скорость обратного движения в четыре раза меньше скорости поступательного движения. Некоторые перитрихи могут перемещаться по поверхности агара, то есть плавающие бактерии способны к передвижению по поверхности твёрдых сред. В частности, Proteus vulgaris распространяется по поверхности агара, образуя тонкий налёт (напоминающий таковой при выдохе на холодное стекло), а неподвижные штаммы протея лишены такой способности. Это явление получило название «феномен роения », а наблюдение за ним легло в основу некоторых понятий бактериальной серодиагностики. Так, жгутиковые Аг называют Н-Аг [от нем. Hauch, выдох, налёт], а Аг клеточной поверхности - О-Аг [от нем. фпе Hauch, без налёта1.

Рис. 4-2. Схема строения бактериального жгутика . БС - базальная структура, ВМ - внешняя мембрана, ЦПМ - цитоплазматическая мембрана, Р - ротор, О - ось, КО - кольцо жгутикового мотора, КР - крюк, С - цилиндрики-соединители, Н - нить жгутика, Ш - шапочка.
Способность бактерий к целенаправленному движению генетически обусловлена. Например, у Escherichia coli в регуляцию этого процесса вовлечено 3% генома (приблизительно 50 генов). Эти гены кодируют белки, образующие локомоторный аппарат, а также белки и ферменты, участвующие в преобразовании сигналов. Для жгутикового аппарата характерна периодическая изменчивость. Во многом этот процесс носит адаптивный характер и наиболее выражен у патогенных микроорганизмов. В частности, некоторые бактерии выработали систему вариабельности антигенных характеристик жгутиков, позволяющую им на какое-то время избегать направленных эффектов защитных иммунных механизмов.
Лабораторная диагностика подвижности бактерий
Подвижность бактерий определяют микроскопией препаратов в «раздавленной » или «висячей » капле. Способность к движению можно определять также после внесения культуры бактерий уколом в столбик полужидкого агара (подвижные виды растут по всей толще среды, неподвижные - по уколу) или посевом бактерий в водный конденсат скошенного столбика агара (подвижные виды переплывают из конденсата на поверхность среды и колонизируют её), либо определяют способность бактерий давать «феномен роения ».
Подвижность очень многих протистов обеспечивается наличием жгутиков или ресничек. И те и другие устроены одинаково. Жгутики и у простейших, и у жгутиковых или ресничных клеток многоклеточных животных и растений всегда лишь часть локомоторной системы клетки, которая состоит из кинетосомы (или центриоли), жгутика (или ундулиподия) и корневых выростов кинетосомы (или ее дериватов (производных)). Кроме передвижения в толще воды, жгутики и реснички применяются для временного или постоянного прикрепления к субстрату или для создания пищевых потоков воды при питании взвешенными частицами.
Жгутик – это трубчатый вырост клеточной поверхности, окруженный мембраной, которая служит продолжением мембраны, покрывающей всю клетку. Он содержит пучок белковых фибрилл, так называемую аксонему. Аксонемой или аксиальной нитью называют микротрубочковое образование, которое состоит из двух центральных микротрубочек, окруженных кольцом из девяти пар (дублетов) микротрубочек, состоящих из плотно спаянных друг с другом субфибрилл. Тонкое строение жгутиков всех эукариотных организмов удивительно однотипно в своих главных чертах.
Важнейшим элементом жгутиковой системы служит базальное тело или кинетосома. Это цилиндр, стенки которого образованы девятью группами микротрубочек, объединенными по три (триплеты). Чаще всего в клетке присутствуют две кинетосомы, расположенные примерно под прямым углом друг к другу. От них отходят один или два жгутика. Кинетосома не плавает в цитоплазме сама по себе, поскольку закреплена системой корешков.
Современные представления о системе Protista во многом базируются на строении жгутика и его производных. Широкое распространение у них жгутиков и ресничек позволяет сравнивать между собой почти все таксоны, а также дает возможность использовать дополнительные признаки жгутикового аппарата, число которых приближается уже к 100, в систематике и филогении. Многие черты строения жгутиконосцев, в том числе и форма тела, определяются наличием у них этой своеобразной системы.
Количество жгутиков, их относительная и абсолютная длина, место и способ прикрепления жгутиков, характер их движения, их направленность очень разнообразны у разных групп, но постоянны внутри отдельных групп родственных организмов.
Обычно выделяют 4 морфотипа жгутиконосцев.
Изоконты имеют от 2 до 8 жгутиков равной длины, направленных в одну сторону, с одинаковыми способами биения. К ним относится большинство подвижных клеток зеленых водорослей.
Анизоконты имеют 2 жгутика неравной длины, направленные в одну сторону, отличающиеся по способу биения. Такие жгутики характерны для бесцветных жгутиконосцев.
У гетероконтных имеются 2 жгутика неравной длины (один направлен вперед, другой - назад), различающиеся по способу биения. Они характерны и для подвижных клеток водорослей, и для так называемых зооспоровых грибов, и для бесцветных жгутиконосцев.
Стефаноконты имеют венчик жгутиков на переднем конце клетки. Это характерно для многожгутиковых гамет и зооспор некоторых зеленых водорослей.
Одножгутиковые формы обычно не выделяются в особую группу. Многие из них рассматриваются как утратившие вторично жгутик особи, так как у подавляющего большинства есть еще одна безжгутиковая кинетосома.
Основная функция жгутика - движение. В активной работе жгутика движущим началом являются периферические микротрубочки и их ручки, обладающие АТФ-азной активностью. Центральные микротрубочки имеют опорное значение. Формы движения жгутика различны, но обычно это винтообразное движение, позволяющее жгутиконосцу «ввинчиваться» в воду, совершая до 40 оборотов в секунду. У инфузорий и многожгутиковых протистов движение ресничек организовано по типу метахрональных волн. Жгутики и реснички часто используются также и для питания. Среди жгутиконосцев имеются виды, проводящие большую часть жизненного цикла в прикрепленном состоянии. В этот период жгутик теряет обычную для него функцию движения и превращается в органеллу прикрепления, стебелек или ножку. Еще одна функция жгутика заключается в том, что он своими движениями очищает поверхность тела от налипающих на него мелких посторонних частиц.
В эндоплазме протистов находятся одно или несколько ядер, а также все органеллы и структуры, свойственные эукариотической клетке: ЭПР, рибосомы, аппарат Гольджи, митохондрии, пероксисомы, гидрогеносомы, пластиды (у автотрофных протистов), лизосомы, пищеварительные вакуоли. Некоторые протисты обладают также специфичными только для них органеллами.
Экструсомы. Эти органеллы представляют собой специальные вакуоли, окруженные мембраной, которая у зрелых экструсом обычно контактирует с плазмалеммой. В ответ на различные внешние раздражения (механические, химические, электрические и др.) они выбрасывают наружу свое содержимое. По своему строению это мукополисахариды (сложные соединения углеводов с белками). Известно 10 разных типов экструсом. Одни содержат ядовитые вещества, способные обездвиживать и убивать жертву (простейших и другие мелкие организмы). Другие выполняют защитную функцию или облегчают движение, выделяя слизь.
Пластиды. Пластиды имеются у фототрофных и близких к ним протистов и представлены хлоропластами и лейкопластами. Основными пигментами хлоропластов являются хлорофиллы. Для разных групп фототрофных протистов характерны определенные наборы хлорофиллов. Из вторичных пигментов у водорослей встречаются каротины и ксантофиллы, которые в больших концентрациях могут маскировать зеленый хлорофилл и придавать хлоропластам разнообразную окраску от желто-зеленой до красновато-коричневой.
Аппарат Гольджи обнаружен практически у всех исследованных видов протистов. Наиболее часто аппарат Гольджи расположен по соседству с ядром и представлен одной или несколькими стопками плоских цистерн (диктиосом), окруженных мелкими пузырьками. Однако аппарат Гольджи у некоторых протистов он образован одиночными цистернами. Отсутствие диктиосом обычно трактуется как примитивный признак. Однако отсутствие диктиосом у современных протистов не может однозначно свидетельствовать об их примитивности, поскольку формирование и разборка диктиосом в большой степени зависят от внешних воздействий на клетку (например, снижение концентрации кислорода в среде) или от физиологических перестроек самого протиста (переход к инцистированию).
Лизосомы и другие органеллы и включения. В клетках протистов, как и в клетках многоклеточных животных, присутствуют лизосомы. Эти цитоплазматические тельца в форме мелких пузырьков (первичные лизосомы) образуются в аппарате Гольджи. В них локализованы пищеварительные гидролитические ферменты. Вторичные лизосомы, или пищеварительные вакуоли, хорошо выражены только у гетеротрофных протистов, питающихся путем фагоцитоза.
В эндоплазме разных протистов в большем или меньшем количестве присутствуют резервные питательные вещества, используемые в процессах метаболизма. Чаще всего это различные полисахариды (гликоген, крахмал, амилоплектин и др.), нередко - липиды и иные жировые включения. Количество резервных веществ зависит от физиологического состояния простейшего, характера и количества пищи, от стадии жизненного цикла и варьирует в широких пределах. Однако некоторые крупные группы протистов запасают специфические вещества. Например, эвгленовые запасают парамил, который не встречается у других протистов.
От переднего полюса тела берут начало жгутики (1, 2, 4, 8 и более - до нескольких тысяч). Если их много, они могут покрывать все тело простейшего (например, в отряде Hypermastigina и отряде Opalinina), напоминая тем самым инфузорий . Длина жгутиков варьирует в широких пределах - от немногих до нескольких десятков микрометров. Если жгутов два, то нередко один выполняет локомоторную функцию, а второй неподвижно тянется вдоль тела и выполняет функцию руля. У некоторых жгутиконосцев (род Trichomonas, род Trypanosoma) жгутик проходит вдоль тела ( рис. 19) и соединяется с последним при помощи тонкой цитоплазматической перепонки. Таким образом формируется ундулирующая мембрана, которая волнообразными колебаниями вызывает поступательное движение простейшего.
В деталях механизм работы жгутиков различен, но в основе это винтообразное движение. Простейшее как бы "ввинчивается" в окружающую среду. Жгутик совершает от 10 до 40 об/с.
Ультраструктура жгутиков очень сложна и обнаруживает поразительное постоянство во всем животном и растительном мире. Все жгутики и реснички животных и растений построены по единому плану (за единичными отклонениями) ( табл. I).
Каждый жгутик слагается из двух отделов. Большую часть его составляет свободный участок, отходящий от поверхности клетки наружу и являющийся собственно локомоторным. Второй отдел жгутика - базальное тело (кинетосома) - меньшая по размерам часть, погруженная в толщу эктоплазмы. Снаружи жгутик покрыт трехслойной мембраной, представляющей непосредственное продолжение наружной мембраны клетки.
Внутри жгутика строго закономерно располагаются 11 фибрилл. Вдоль оси жгута проходят 2 центральные фибриллы ( рис. 20), берущие начало от аксиальной гранулы. Диаметр каждой из них составляет около 25 нм, а центры их расположены на расстоянии 30 нм. По периферии под оболочкой находятся еще 9 фибрилл, причем каждая слагается из двух тесно спаянных трубочек. Локомоторная деятельность жгутика определяется периферическими фибриллами, тогда как центральные играют опорную функцию и может быть представляют собой субстрат, по которому распространяются волны возбуждения, вызывающие движение жгутика.
Базальное тело (кинетосома) , находится в эктоплазме. Оно имеет вид цилиндрического тельца, окруженного мембраной, под которой по периферии располагаются 9 фибрилл, являющихся непосредственным продолжением периферических фибрилл самого жгута. Здесь они, правда, становятся тройными ( рис. 20 , табл. II). Иногда основание жгутика продолжается в глубину цитоплазмы за пределы кинетосомы, образуя корневую нить (ризопласт), которая может либо свободно заканчиваться в цитоплазме, либо прикрепляться к оболочке ядра.
У некоторых жгутиконосцев вблизи кинетосомы располагается парабазальное тело . Форма его может быть разнообразной. Иногда это яйцевидное или колбасовидное образование, иногда же оно приобретает довольно сложную конфигурацию и состоит из множества отдельных долек (
Жгутики прокариот
Жгутикование бактерий: A - монотрихиальное,
B - лофотрихиальное,
C - амфитрихиальное,
D - перитрихиальное.
Ссылки
Литература
Wikimedia Foundation . 2010 .
Смотреть что такое "Жгутики" в других словарях:
Нитевидные подвижные цитоплазматические выросты клетки, свойственные многим бактериям, всем жгутиковым, зооспорам и сперматозоидам животных и растений. Служат для передвижения в жидкой среде … Большой Энциклопедический словарь
Органоиды движения бактерий, представляют собой тонкие длинные (3 5x12 25 нм) нитевидные структуры белковой природы. Начинаются в цитоплазме от базального тельца. Далее они, окруженные футляром, утолщенные и изогнутые в виде крюка, проходят через … Словарь микробиологии
жгутики - реснички Волосоподобные выросты поверхностных клеток небольших организмов, с помощью которых эти организмы передвигаются или пропускают воду сквозь свое тело; обнаружены в механизмах питания многих морских животных.… … Справочник технического переводчика
Нитевидные подвижные цитоплазматические выросты клетки, свойственные многим бактериям, всем жгутиковым, зооспорам и сперматозоидам животных и растений. Служат для передвижения в жидкой среде. * * * ЖГУТИКИ ЖГУТИКИ, нитевидные подвижные… … Энциклопедический словарь
Цитоплазматические выросты клетки, свойственные всем жгутиковым (См. Жгутиковые), многим бактериям, зооспорам и сперматозоидам растений и животных. Раньше Ж. противопоставляли ресничкам (См. Реснички). Электронномикроскопические… … Большая советская энциклопедия
Нитевидные подвижные цитоплазматич. выросты клетки, свойственные мн. бактериям, всем жгутиковым, зооспорам и сперматозоидам ж ных и р ний. Служат для передвижения в жидкой среде … Естествознание. Энциклопедический словарь
жгутики - длинные и относительно толстые клеточные структуры, в образовании которых принимают участие плазмалемма и микротрубочки; встречаются у одноклеточных и колониальных водорослей: хламидомонады (Chlamydomonas), вольвокса (Volvox), a также у зооспор,… … Анатомия и морфология растений
Как прокариотические, так и эукариотические могут содержат структуры, известные как реснички и жгутики. Эти выросты на поверхности клеток помогают в их .
Особенности и функции
Реснички и жгутики являются выростами из некоторых клеток необходимые для клеточной локомоции (передвижения). Они также помогают перемещать вещества вокруг клеток и направлять их к нужным участкам.
Реснички и жгутики образуются из специализированных групп микротрубочек, называемых базальными телами.
Если выросты короткие и многочисленные, их называют ресничками. Если они длиннее и менее многочисленны (обычно только один или два), они называются жгутиками.
Строение
Обычно реснички и жгутики имеют сердцевину, состоящую из микротрубочек, соединенных с , расположенных по схеме 9+2. Кольцо из девяти микротрубочек имеет в своем центре две особые микротрубочки, которые сгибают реснички или жгутики. Этот тип организации встречается в устройстве большинства ресничек и жгутиков .
Где встречаются?
Как реснички, так и жгутики встречаются во многих типах клеток. Например, у спермы многих животных, водорослей и даже папоротников есть жгутики. Реснички можно найти в клетках таких тканей, как дыхательные пути и женский репродуктивный тракт.
Цитоплазма бактерий. Бактериальный геном. Бактериальные рибосомы. Запасные гранулы бактерии.
Структура бактериальной клетки
Бактериальная клетка состоит из клеточной стенки, цитоплазматической мембраны, цитоплазмы с включениями и ядерного аппарата, называемого нуклеоидом. Имеются другие структуры: мезосома, хроматофоры, тилакоиды, вакуоли, включения полисахаридов, жировые капельки, капсула (микрокапсула, слизь), жгутики, пили. Некоторые бактерии способны образовывать споры.
Структуру и морфологию бактерий изучают с помощью различных методов микроскопии: световой, фазово-контрастной, интерференционной, темнопольной, люминесцентной и электронной.
Обозначения:
Клеточная стенка
В клеточной стенки грамположительных бактерий содержится небольшое количество полисахаридов, липидов, белков. Основным компонентом клеточной стенки этих бактерий является многослойный пептидогликан (муреин, мукопептид), составляющий 40—90% массы клеточной стенки. С пептидогликаном клеточной стенки грамположительных бактерий ковалентно связаны тейхоевые кислоты (от греч. teichos — стенка).
В состав клеточной стенки грамотрицательных бактерий входит наружная мембрана, связанная посредством липопротеина с подлежащим слоем пептидогликана. На ультратонких срезах бактерий наружная мембрана имеет вид волнообразной трехслойной структуры, сходной с внутренней мембраной, которую называют цитоплазматической. Основным компонентом этих мембран является бимолекулярный (двойной) слой липидов. Внутренний слой наружной мембраны представлен фосфолипидами, а в наружном слое расположен липополисахарид (ЛПС). Липополисахарид наружной мембраны состоит из трех фрагментов: липида А - консервативной структуры, практически одинаковой у грамотрицательных бактерий; ядра, или стержневой, коровой части (лат. core — ядро), относительно консервативной олигосахаридной структуры (наиболее постоянной частью ядра ЛПС является кетодезоксиоктоновая кислота); высоковариабельнои О-специфической цепи полисахарида, образованной повторяющимися идентичными олигосахаридными последовательностями (О-антиген). Белки матрикса наружной мембраны пронизывают ее таким образом, что молекулы белка, называемые поринами, окаймляют гидрофильные поры, через которые проходят вода и мелкие гидрофильные молекулы.
При нарушении синтеза клеточной стенки бактерий под влиянием лизоцима,
пенициллина, защитных факторов организма образуются клетки с измененной (часто шаровидной) формой: протопласты — бактерии, полностью лишенные клеточной стенки; сферопласты - бактерии с частично сохранившейся клеточной стенкой. Бактерии сферо- или протопластного типа, утратившие способность к синтезу пептидогликана под влиянием антибиотиков или других факторов и способные размножаться, называются L-формами.
Они представляют собой осмотически чувствительные, шаровидные, колбовидные клетки различной величины, в том числе и проходящие через бактериальные фильтры. Некоторые L-формы (нестабильные) при удалении фактора, приведшего к изменениям бактерий, могут реверсировать, «возвращаясь» в исходную бактериальную клетку.
Между наружной и цитоплазматической мембранами находится периплазматическое пространство, или периплазма, содержащая ферменты (протеазы, липазы, фосфатазы, нуклеазы, бета-лактомазы) и компоненты транспортных систем.
Цитоплазматическая мембрана
Цитоплазматическая мембрана при электронной микроскопии ультратонких срезов представляет собой трехслойную мембрану (2 темных слоя толщиной по 2,5 нм разделены светлым - промежуточным). По структуре она похожа на плазмалемму клеток животных и состоит из двойного слоя фосфолипидов с внедренными поверхностными, а также интегральными белками, как бы пронизывающими насквозь структуру мембраны. При избыточном росте (по сравнению с ростом клеточной стенки) цитоплазматическая мембрана образует инвагинаты — впячивания в виде сложно закрученных мембранных структур, называемые мезосомами. Менее сложно закрученные структуры называются внутрицитоплазматическими мембранами.
Цитоплазма
Цитоплазма состоит из растворимых белков, рибонуклеиновых кислот, включений и многочисленных мелких гранул — рибосом, ответственных за синтез (трансляцию) белков. Рибосомы бактерий имеют размер около 20 нм и коэффициент седиментации 70S, в отличие от 80S-рибосом, характерных для эукариотических клеток. Рибосомные РНК (рРНК) - консервативные элементы бактерий («молекулярные часы» эволюции). 16S рРНК входит в состав малой субъединицы рибосом, а 23S рРНК - в состав большой субъединицы рибосом. Изучение 16S рРНК является основой геносистематики, позволяя оценить степень родства организмов.
В цитоплазме имеются различные включения в виде гранул гликогена, полисахаридов, бета-оксимасляной кислоты и полифосфатов (волютин). Они являются запасными веществами для питания и энергетических потребностей бактерий. Волютин обладает сродством к основным красителям и легко выявляется с помощью специальных методов окраски (например, по Нейссеру) в виде метахроматических гранул. Характерное расположение гранул волютина выявляется у дифтерийной палочки в виде интенсивно прокрашивающихся полюсов клетки.
Нуклеоид
Нуклеоид — эквивалент ядра у бактерий. Он расположен в центральной зоне бактерий в виде двунитевой ДНК, замкнутой в кольцо и плотно уложенной наподобие клубка. Ядро бактерий, в отличие от эукариот, не имеет ядерной оболочки, ядрышка и основных белков (гистонов). Обычно в бактериальной клетке содержится одна хромосома, представленная замкнутой в кольцо молекулой ДНК.
Кроме нуклеоида, представленного одной хромосомой, в бактериальной клетке имеются внехромосомные факторы наследственности - плазмиды, представляющие собой ковалентно замкнутые кольца ДНК.
Капсула, микрокапсула, слизь
Капсула - слизистая структура толщиной более 0,2мкм, прочно связанная с клеточной стенкой бактерий и имеющая четко очерченные внешние границы. Капсула различима в мазках-отпечатках из патологического материала. В чистых культурах бактерий капсула образуется реже. Она выявляется при специальных методах окраски мазка (например, по Бурри-Гинсу), создающих негативное контрастирование веществ капсулы: тушь создает темный фон вокруг капсулы. Капсула состоит из полисахаридов (экзополисахаридов), иногда из полипептидов, например, у сибиреязвенной бациллы она состоит из полимеров D-глутаминовой кислоты. Капсула гидрофильна, препятствует фагоцитозу бактерий. Капсула антигенна: антитела против капсулы вызывают ее увеличение (реакция набухания капсулы).
Многие бактерии образуют микрокапсулу - слизистое образование толщиной менее 0,2мкм, выявляемое лишь при электронной микроскопии. От капсулы следует отличать слиэь - мукоидные экзополисахариды, не имеющие четких границ. Слизь растворима в воде.
Бактериальные экзополисахариды участвуют в адгезии (прилипании к субстратам), их еще называют гликокаликсом. Кроме синтеза
экзополисахаридов бактериями, существует и другой механизм их образования: путем действия внеклеточных ферментов бактерий на дисахариды. В результате этого образуются декстраны и леваны.
Жгутики
Жгутики бактерий определяют подвижность бактериальной клетки. Жгутики представляют собой тонкие нити, берущие начало от цитоплазматической мембраны, имеют большую длину, чем сама клетка. Толщина жгутиков 12-20 нм, длина 3-15 мкм. Они состоят из 3 частей: спиралевидной нити, крюка и базального тельца, содержащего стержень со специальными дисками (1 пара дисков - у грамположительных и 2 пары дисков - у грамотрицательных бактерий). Дисками жгутики прикреплены к цитоплазматической мембране и клеточной стенке. При этом создается эффект электромотора со стержнем-мотором, вращающим жгутик. Жгутики состоят из белка - флагеллина (от flagellum - жгутик); является Н-антигеном. Субъединицы флагеллина закручены в виде спирали.
Число жгутиков у бактерий различных видов варьирует от одного (монотрих) у холерного вибриона до десятка и сотен жгутиков, отходящих по периметру бактерии (перитрих) у кишечной палочки, протея и др. Лофотрихи имеют пучок жгутиков на одном из концов клетки. Амфитрихи имеют по одному жгутику или пучку жгутиков на противоположных концах клетки.
Пили (фимбрии, ворсинки) - нитевидные образования, более тонкие и короткие (3-10нм х 0, 3-10мкм) , чем жгутики. Пили отходят от поверхности клетки и состоят из белка пилина, обладающего антигенной активностью. Различают пили, ответственные за адгезию, то есть за прикрепление бактерий к поражаемой клетке, а также пили, ответственные за питание, водносолевой обмен и половые (F-пили), или конъюгационные пили. Пили многочисленны - несколько сотен на клетку. Однако, половых пилей обычно бывает 1-3 на клетку: они образуются так называемыми "мужскими" клетками-донорами, содержащими трансмиссивные плазмиды (F-, R-, Col-плазмиды). Отличительной особенностью половых пилей является взаимодействие с особыми "мужскими" сферическими бактериофагами, которые интенсивно адсорбируются на половых пилях.
Споры
Споры - своебразная форма покоящихся фирмикутных бактерий, т.е. бактерий
с грамположительным типом строения клеточной стенки. Споры образуются при неблагоприятных условиях существования бактерий (высушивание, дефицит питательных веществ и др.. Внутри бактериальной клетки образуется одна спора (эндоспора). Образование спор способствует сохранению вида и не является способом размножения, как у грибов. Спорообразующие бактерии рода Bacillus имеют споры, не превышающие диаметр клетки. Бактерии, у которых размер споры превышает диаметр клетки, называются клостридиями, например, бактерии рода Clostridium (лат. Clostridium - веретено). Споры кислотоустойчивы, поэтому окрашиваются по методу Ауески или по методу Циля-Нильсена в красный, а вегетативная клетка в синий цвет.
Форма спор может быть овальной, шаровидной; расположение в клетке -терминальное, т.е. на конце палочки (у возбудителя столбняка), субтерминальное - ближе к концу палочки (у возбудителей ботулиэма, газовой гангрены) и центральное (у сибиреязвенной бациллы). Спора долго сохраняется из-за наличия многослойной оболочки, дипиколината кальция, низкого содержания воды и вялых процессов метаболизмов. В благоприятных условиях споры прорастают, проходя три последовательные стадии: активация, инициация, прорастание.
Цитоплазма бактерий. Бактериальный геном. Бактериальные рибосомы. Запасные гранулы бактерии.
Цитоплазма бактерий. Бактериальный геном. Бактериальные рибосомы. Запасные гранулы бактерии.
Цитоплазма бактерий представляет собой коллоидный матрикс, служащий для реализации жизненно важных функций. Цитоплазма большинства бактерий содержит ДНК, рибосомы и запасные гранулы; остальное пространство занимает коллоидная фаза. Её основные составляющие — растворимые ферменты и растворимые РНК (мРНК и тРНК). Разнообразные органеллы, характерные для эукариотической клетки, у бактерий отсутствуют, а их функции выполняет бактериальная ЦПМ, отделяющая цитоплазму от клеточной стенки. У подавляющего числа бактерий цитоплазма относительно неподвижна, но у видов Streptococcus, Proteus, Clostridium имеются специальные трубочки — рапидосомы, аналогичные микротрубочкам простейших.
Бактериальный геном
В бактериальной клетке нет ядерной мембраны, ДНК сконцентрирована в цитоплазме в виде клубка. Поскольку в эукариотических клетках ДНК обычно находится в ядре, то по аналогии ДНК бактерий назвали нуклеоидом [от лат. nucleus, ядро + греч. eidos, сходство]. Её также называют генофором, или бактериальной хромосомой. Генофор бактерий представлен двойной спиральной, кольцевой, ковалентно замкнутой суперспирализованной молекулой ДНК. Она составляет 2-3% сухой массы клетки (более 10% по объёму). Генофор не содержит гистонов. Объём генетической информации, кодируемой в генофоре, различается в зависимости от вида бактерии (например, геном Escherichia coli кодирует примерно 4000 различных полипептидов). У бактерий может присутствовать дополнительная ДНК в виде включений. Эти включения, или плазмиды, несут ряд различных генов, кодирующих дополнительные свойства бактерий, но информация, содержащаяся в плазмидах, не является абсолютно необходимой для бактериальной клетки.

Бактериальные рибосомы
Бактериальные рибосомы — сложные глобулярные образования, состоящие из различных молекул РНК и связанных с ними белков. Всё образование функционирует как локус синтеза полипептидов. В зависимости от интенсивности роста бактериальная клетка может содержать от 5000 до 50 000 рибосом. Диаметр бактериальных рибосом около 16-20 нм. Скорость их осаждения при ультрацентрифугировании составляет 70 S (единиц Свёдберга), тогда как у эукариотических клеток — 80 S. Рибосомы бактерий состоят из двух субъединиц с коэффициентом седиментации 50 S и 30 S (у эукариотов 40 S и 60 S). Объединение субъединиц происходит перед началом трансляции. Рибосомы прокариот и эукариотов имеют сходную молекулярную структуру и механизмы функционирования, но различаются, помимо размеров, по составу белков и белковых факторов. Эти различия делают рибосомы эукариотов практически резистентными к действию антибиотиков, блокирующих синтез белка у бактерий.
Запасные гранулы бактерии
Запасные гранулы содержат временный избыток метаболитов; наличие и количество гранул изменяется в зависимости от вида бактерий и их метаболической активности. В виде гранул могут запасаться полисахариды (крахмал, гликоген, гранулёза), жиры (триглицериды, сходные с жирами высших животных, запасаются у дрожжей рода Candida), воска — у микобактерий и нокардий; полимеры р-оксимасляной кислоты (например, в клетках Bacillus megaterium), полифосфаты (волютин) у Spirillum volutans и Corynebacterium diphtheriae, сера (у бактерий, окисляющих сульфиды), кристаллизованные белки (например, токсичный для насекомых протоксин у Bacillus thuringiensis).
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Цитоплазмамическая мембрана

В состав клеточной стенки грамотрицательных бактерий входит наружная мембрана, связанная посредством липопротеина с подлежащим слоем пептидогликана. На ультратонких срезах бактерий наружная мембрана имеет вид волнообразной трехслойной структуры, сходной с внутренней мембраной, которую называют цитоплазматической. Основным компонентом этих мембран является бимолекулярный (двойной) слой липидов. Внутренний слой наружной мембраны представлен фосфолипидами, а в наружном слое расположен липополисахарид. Липополисахарид наружной мембраны состоит из 3 фрагментов: липида А — консервативной структуры, практически одинаковой у грамотрицательных бактерий; ядра, или стержневой, коровой части (от лат. core— ядро), относительно консервативной олигосахаридной структуры (наиболее постоянной частью ядра ЛПС является кетодезоксиоктоновая кислота); высоковариабельной О-специфической цепи полисахарида, образованной повторяющимися идентичными олигосахаридными последовательностями (О-антиген). Белки матрикса наружной мембраны пронизывают ее таким образом, что молекулы белка, называемые поринами, окаймляют гидрофильные поры, через которые проходят вода и мелкие гидрофильные молекулы.
При нарушении синтеза клеточной стенки бактерий под влиянием различных факторов образуются клетки с измененной формой (чаще шаровидной): протопласты— бактерии, полностью лишенные клеточной стенки;сферопласты— бактерии с частично сохранившейся клеточной стенкой. Бактерии сферо- или протопластного типа, утратившие способность к синтезу пептидогликана и способные размножаться, называютсяL-формами. Существуют нестабильные L-формы, которые могут реверсировать, «возвращаясь» в исходную бактериальную клетку.и стабильные L-формы, не способные к реверсии.
Цитоплазматическая мембрана. ЦПМ - это обязательная клеточная структура, являющаяся физическим, осмотическим, метаболическим барьером между внутренним содержимым бактериальной клетки и внешней средой. ЦПМ состоит из двух слоев фосфолипидов и встроенных в липидную мембрану белковых молекул (как большинство биологических мембран). Белки составляют 20-75%, липиды – 25-40% и в незначительных количествах в ЦПМ присутствуют углеводы и РНК, Белки ЦПМ подразделяют на структурные и функциональные. Первые образуют различные структуры ЦПМ, вторые представлены ферментами, участвующими в синтетических реакциях на поверхности мембраны и окислительно-восстановительных процессах, а также некоторыми специализированными энзимами (пермеазы участвуют в транспорте веществ). При избыточном росте (по сравнению с ростом клеточной стенки) цитоплазматическая мембрана образует инвагинаты — впячивания в виде сложно закрученных мембранных структур, называемые мезосомами. Мезосомы образуют поперечные перегородки между делящимися клетками и являются местом прикрепления бактериальной хромосомы.
У некоторых бактерий между ЦПМ и клеточной стенкой располагается периплазматическое пространство – полость шириной около 10 нм. Снаружи в это пространство открываются поры клеточной стенки, изнутри выходят некоторые клеточные ферменты (рибонуклеазы, фосфатазы, β-лактамазы).
Цитоплазма. Цитоплазма бактерий представляет собой коллоидный матрикс, служащий для реализации жизненно важных функций. Цитоплазма большинства бактерий содержит ДНК, рибосомы и запасные гранулы; остальное пространство занимает коллоидная фаза. Ее основные составляющие – растворимые ферменты и РНК (мРНК и тРНК).
Бактериальный геном.Эквивалентом ядра у бактерий является нуклеоид (генофор). Он расположен в центральной зоне бактерий в виде двойной суперспирализованной кольцевой молекулой ДНК. Она составляет 2-3% сухой массы клетки (более 10% по объему). Генофор не содержит гистонов. Объем генетической информации, кодируемой в бактериальной хромосоме зависит от вида бактерии. Кроме нуклеоида в бактериальной клетке имеются внехромосомные носители генетической информации – плазмиды, являющиеся ковалентно замкнутыми кольцами ДНК. Плазмиды несут ряд различных генов, кодирующих дополнительные (необязательные) признаки бактерий, например, гены антибиотикорезистентности (R-фактор). Бактерии могут обмениваться плазмидами в процессе конъюгации.
Рибосомы. Бактериальные рибосомы – сложные глобулярные образования, состоящие из молекул РНК и связанных с ними белками. Рибосомы необходимы для синтеза полипептидов. Количество рибосом в различных бактериальных клетках колеблется от 5 до 50 тысяч. Диаметр рибосом составляет около 16-20 нм. Скорость их осаждения при ультрацентрифугировании составляет 70S(единиц Сведберга), тогда как у эукариотических клеток 80S. Рибосомы бактерий состоят из двух субъединиц с коэффициентом седиментации 50Sи 30S(у эукариот 40Sи 60S). Объединение субъединиц происходит перед началом трансляции. Рибосомальные РНК (рРНК) — консервативные элементы бактерий («молекулярные часы» эволюции). 16S рРНК входит в состав малой субьединицы рибосом, а 23S рРНК — в состав большой субъединицы рибосом. Изучение 16S рРНК является основой геносистематики, позволяющей оценить степень родства организмов. Различия в строении рибосом про- и эукариотических клеток делают рибосомы эукариот практически резистентными к действию антибиотиков, блокирующих синтез белка у бактерий.
Запасные гранулы.В цитоплазме бактерий содержатся различные включения, которые содержат временный избыток метаболитов. В виде гранул могут запасаться полисахариды (крахмал, гликоген), жиры (триглицериды, запасаются у дрожжеподобных грибов родаCandida), полимеры β-оксимасляной кислоты, полифосфаты (волютин) уC.diphtheriae, сера, кристаллизованные белки и др.
Жгутики. Жгутики бактерий являются органами движения (локомоции) бактерий. Расположение жгутиков – характерный признак, имеющий таксономическое значение. По количеству и расположению жгутиков различают монотрихи – один жгутик (V.cholerae), перитрихи (от греч.peri, вокруг +trichos, волос)– жгутики по всей поверхности бактериальной клетки (E.coli), лофотрихи (от греч.lophos, пучок +trichos, волос)– пучок жгутиков на одном конце клетки (Pseudomonas), амфитрихи (от греч.amphi, двойной, двусторонний +trichos, волос) –единичные жгутики или пучки жгутиков на разных полюсах клетки (Spirillum). Жгутик – спирально изогнутая полая нить, образованная субъединицами белка флагеллина. Толщина жгутиков 12-20 нм, длина 3-15 мкм (больше длины клетки). Жгутики состоят из 3 частей: спиралевидной нити, крюка и базального тельца. Базальное тельце содержит стержень со специальными дисками: одна пара дисков — у грамположительных и 2 пары дисков — у грамотрицательных бактерий. Дисками жгутики прикреплены к цитоплазматической мембране и клеточной стенке. При этом создается эффект электромотора со стержнем-мотором, вращающим жгутик. Бактериальные жгутики совершают поступательные и вращательные движения, проталкивая бактерии через среду подобно корабельному винту. Жгутики являютсяH-антигенами, что используется в серологической идентификации.
Подвижность бактерий определяют микроскопией препаратов «раздавленной» или «висячей» капли. Способность к движению можно определять также после внесения культуры бактерий уколом в столбик полужидкого агара (подвижные виды растут по всей толще среды, неподвижные – по уколу) или посевом бактерий в водный конденсат скошенного столбика агара (подвижные виды переплывают из конденсата на поверхность среды и колонизируют ее).
Микроворсинки.Помимо жгутиков, поверхность многих бактерий покрыта цитоплазматическими выростами – микроворсинками, встречающимися у подвижных и неподвижных видов. Пили (фимбрии, ворсинки) — нитевидные образования, более тонкие и короткие (3-10 нм х 0,3-10 мкм), чем жгутики. Пили отходят от поверхности клетки и состоят из белка пилина, обладающего антигенной активностью. Различают пили, ответственные за адгезию, прикрепление бактерий к поражаемой клетке; пили, ответственные за питание, водно-солевой обмен и половые (F-пили), или коньюгационные, пили. Пили многочисленны — несколько сотен на клетку. Однако половых пилей обычно бывает 1-3 на клетку: они образуются так называемыми «мужскими» клетками-донорами, содержащими трансмиссивные плазмиды (F-, R-, Col- плазмиды). Отличительной особенностью половых пилей является взаимодействие с особыми «мужскими» сферическими бактериофагами, которые интенсивно адсорбируются на половых пилях.
Споры.Споры образуются при неблагоприятных условиях существования бактерий (высушивание, дефицит питательных веществ и др.). Внутри бактериальной клетки образуется одна спора (эндоспора). Образование спор способствует сохранению вида и не является способом размножения, как у грибов. Способностью к спорообразованию обладают только бактерии родовBacillusиClostridium, включающие патогенные для человека виды, и многие сапрофиты. У бактерий родаBacillusдиаметр споры не превышает диаметр (поперечник) бактериальной клетки. У бактерий родаClostridium(от лат.closter, веретено) диаметр споры превышает поперечник бактерии, что придает им форму веретена (C.perfringens), барабанной палочки (C.tetani) или теннисной ракетки (C.botulinum) при центральной, терминальной и субтерминальной локализации спор соответственно. Основное значение спор в выживаемости бактерий определяет их терморезистентность и резистентность к другим факторам (дезинфектанты и др.), оказывающим губительное воздействие на бактерии. Высокая устойчивость спор связана с низким содержанием свободной воды, высокой концентрацией кальция, вялостью метаболических процессов, наличием дипиколиновой кислоты и белка, богатого цистеином (что делает его похожим на кератин), а также наличие нескольких оболочек, которые являются дополнительной защитой от неблагоприятных внешних воздействий. При попадании спор в благоприятные условия они прорастают, проходя три последовательные стадии: активацию, инициацию, вырастание.
Споры плохо окрашиваются и остаются бесцветными в окрашенных клетках при использовании обычных методов окраски (простые методы, метод Грама). Для окраски спор используют методы Ожешко (Ауэски).
Цитоплазма и включения
Цитоплазма — гетерогенная система, в которой различают дисперсионную среду и дисперсную фазу. Дисперсионной средой является вода с растворенными в ней низкомолекулярными веществами, а дисперсной фазой — высокомолекулярные соединения: белки, липиды, углеводы и их конъюганты. Соотношение свободной и связанной воды неодинаково у разных микроорганизмов и меняется у одного и того же штамма с возрастом, в зависимости от условий выращивания. Ацетон, метанол, этанол легко отнимают свободную воду и воду гидратных оболочек, приводя к коагуляции макромолекулярные структуры.
В цитоплазме одни вещества находятся в виде истинного раствора (неорганические соли, витамины, аминокислоты), а другие — в коллоидном состоянии (белки). Внутриклеточное осмотическое давление, например, бактериальной клетки эквивалентно 10—20%-ным растворам сахарозы. Консистенция цитоплазмы в разных слоях неодинакова: располагающийся непосредственно под ЦПМ тонкий слой более плотный, чем остальная масса в центре клетки. Многие молекулы в цитоплазме заряжены, из них белковые несут положительный и отрицательный заряды одновременно.
Разнообразные органеллы, характерные для эукариотной клетки, у бактерий отсутствуют, а их функции выполняет бактериальная ЦПМ, отделяющая цитоплазму от клеточной стенки. Цитоплазма относительно неподвижна, но у видов Streptococcus, Proteus, Clostridium имеются специальные трубочки — рапидосомы, аналогичные микротрубочкам простейших. Они являются как бы якорной структурой для ядра. Рапидосомы (от лат. rapidus — быстрый), или микротрубочки (микротубули), могут быть в виде полого или неполого цилиндра размером 33 х 225 нм.
Ядерный аппарат, или нуклеоид (от лат. nucleus — ядро + греч. eidos — сходство). Классические цитологические методы, ультра- тонкие срезы в сочетании с электронной микроскопией позволили установить, что бактерии содержат ДНК, которая не распределена диффузно в цитоплазме, а локализована в ограниченных участках, которые делятся перед делением клетки. Область ядра (нуклеоплазма) в бактериальной клетке равномерно заполнена очень тонкими нитями. Ее называют генофором, или бактериальной хромосомой. Генофор бактерий представлен двойной спиральной, кольцевой, ковалентно замкнутой суперспирализованной молекулой ДНК, длина ее контура составляет от 0,25 до 3 мм. Она составляет 2—3% сухой массы клетки (более 10% по объему). Объем генетической информации, кодируемой в генофоре, различается в зависимости от вида бактерий (геном Е. coli, например, кодирует примерно 4000 различных полипептидов). У бактерий может присутствовать дополнительная ДНК в виде включений — плазмид, которые несут ряд различных генов, кодирующих дополнительные свойства бактерий. Информация, содержащаяся в плазмидах, не является абсолютно необходимой для бактериальной клетки. Плазмиды могут существовать в клетке и в интегрированном состоянии с бактериальной хромосомой, сохраняя при этом способность переходить к автономии. Будучи интегрированными, они не являются частью линейной структуры хромосомы. Автономность и интегрированность плазмид исключают друг друга.
Микробные клетки эукариот содержат оформленное ядро, отделенное от цитоплазмы двухслойной мембраной. Химическая природа ядра довольно сложная. Ядро содержит ДНК и основные белки — гистоны. Хромосомный материал в виде компактных гранул, включающий по одной молекуле ДНК на хромосому, погружен в ядерную основу. Молекулярная масса ДНК одной хромосомы (из 17) у дрожжей, например, составляет 6 • 10 5 кДа.
Включения или запасные гранулы. В цитоплазме клеток бактерий в определенных условиях среды откладываются вещества, которые можно рассматривать как запасные, — полисахариды, жиры, полифосфаты и сера. Они содержатся в клетках в осмотически инертной форме и нерастворимы в воде. При условиях, благоприятных для роста бактерий, когда в этих веществах возникает потребность, запасные вещества снова включаются в метаболизм.
Запасные полисахариды, нейтральные жиры и поли-р-гидро- ксимасляная кислота могут служить источниками как энергии, так и углерода. Поэтому в «экстремальных условиях» (отсутствие внешних источников энергии) они могут продлить время существования клетки, а у спорообразующих видов — создать условия для образования спор. Полифосфаты могут служить резервным источником фосфата, а запасная сера (в виде капелек) — как потенциальный донор электронов. Их присутствие нельзя рассматривать как какой-то постоянный признак микроорганизма; в значительной степени это связано с физическими и химическими условиями среды обитания. Запасные гранулы выявляются, как правило, при выращивании микроорганизмов в средах, содержащих все необходимые питательные вещества, а исчезают при голодании клеток. Количество гранул изменяется в зависимости от вида бактерий и их метаболической активности.
В виде гранул могут запасаться полисахариды (крахмал, гликоген, гранулеза); жиры (триглицериды, сходные с жирами высших животных, запасаются у дрожжей рода Candida); воска — у микобактерий и нокардий; полимеры $-оксимасляной кислоты (в клетках Вас. megaterium); полифосфаты (волютин); сера (у бактерий, окисляющих сульфиды); кристаллизованные белки (токсичный для насекомых протоксин у Вас. thuringiensis).
Крахмал — резервное питательное вещество клетки бактерий, обнаруживается при помощи цветной реакции с раствором Лю- голя (синяя окраска). Крахмалоподобное соединение у клостри- дий, обнаруживаемое раствором I2 + KI, получило название гранулеза. «Жидкий крахмал», или гликоген, сходен с амилопектином и обнаруживается у дрожжей, грибов и других микроорганизмов; найден также в тканях животных. По строению он подобен ами- лопектину, но расстояние между точками ветвления почти в два раза меньше (10—14 остатков глюкозы). Гликоген, изолированный из печени крысы, представляется в виде крупных агрегатов диаметром 600—2000 А. Подобные частицы были обнаружены в цитоплазме дрожжей. При недостаточном поступлении углеродсодержащих веществ в питательную среду гранулы гликогена или гранулезы постепенно исчезают из клеток бактерий.
Жиры в клетках микроорганизмов часто встречаются в виде гранул и капелек. В световом микроскопе они видны благодаря сильному преломлению света и окрашиванию липофильным красителем Судан III. Жировые включения ряда бактерий представлены в виде гранул поли-р-гидроксимасляной кислоты, а также другими липидами. Поли-р-гидроксимасляная кислота синтезируется из углеродсодержащих соединений при дефиците источников азота в среде. У факультативных анаэробов и строгих аэробов она накапливается, когда клетки испытывают недостаток кислорода и переходят на брожение; поэтому ее можно рассматривать как полимерный внутриклеточный продукт брожения. В аэробных условиях этот продукт может снова использоваться как источник энергии и углерода в процессе дыхания. Гранулы ее покрыты мембраной, но их не относят к числу органелл. Доля этого вещества в сухой биомассе может достигать 80%.
Сложные эфиры жирных кислот с длинной цепью и спиртов — воска могут накапливаться в клетках до 40% (у микобактерий). Содержание запасных жиров определяется составом питательной среды (высоким отношением C/N). Количества других липидных соединений от состава среды почти не зависят.
Полифосфатсодержащие гранулы (волютин). Волютиновые гранулы, называемые еще метахроматическими гранулами (впервые описаны у Spirillum volutans), состоят преимущественно из полифосфатов, полиметафосфатов, а также соединений, близких к нуклеиновым кислотам. Характерной особенностью волютина является хромофильность и метахромазия, т.е. он обладает ясно выраженным сродством к основным красителям и часто окрашивается в необычный цвет (например, в красный при применении метиленового синего). Образуется в виде крупных, отчетливо видимых гранул в больших количествах на средах, богатых глицерином или углеводами. Как источники энергии полифосфаты играют второстепенную роль.
Кристаллизованные белки — протоксины используются для биологической борьбы с вредителями. Обнаруживаются у Bacillus thuringiensis и родственных видов.
Сера — минеральное включение бактериальной клетки. У серных бактерий сера временно сохраняется в виде полужидких шариков, сильно преломляющих свет. Количество накапливаемой серы зависит от содержания сероводорода в окружающей среде: при его отсутствии сера окисляется до сульфата. Для аэробных серных бактерий (Beggiatoa) сера служит источником энергии, а для анаэробных фототрофных пурпурных серобактерий (Chromatium) — донором электронов. Включения серы можно рассматривать как продукты обезвреживания сероводорода, часто присутствующего в местах обитания цианобактерий. Некоторые серные бактерии наряду с капельками серы имеют зернышки аморфного карбоната кальция, роль которого пока не выяснена.
Спора (от греч. spora — сияние, семя) бактерий — покоящиеся репродуктивные клетки, характеризующиеся резко сниженным уровнем метаболизма и особой ультраструктурной организацией. Основная функция — репродукция и (или) сохранение бактерий в неблагоприятных условиях внешней среды. Эндоспоры — уникальные покоящиеся формы, формирующиеся обычно по одной внутри материнских клеток, устойчивые к высоким температурам, образуют только истинные бактерии из семейства ВасШасеае (роды Bacillus и Clostrdium) (рис. 3.18).
Эндоспоры обладают специфическими структурами (белковыми покровами, пептидогликановым кортексом, сердцевиной). Содержат в отличие от вегетативных клеток дипиколиновую кислоту (пиридин-2,6-дикарбоновую) в виде Са-хелата. Устойчивы к стрессам (высоким температурам, радиации, высушиванию, действию растворителей и др.). Споры бактерий представляют собой овальные или сферические тельца, сильно преломляющие свет, окруженные толстой оболочкой. В сухом состоянии они погибают лишь при нагревании в течение 2 часов при 165°С, а в перегретом паре — при 12ГС за 15 мин. Считается, что для уничтожения спор требуется температура пара 120°С при давлении его в 1,01 • 10 5 Па в течение 20 мин.
Эндоспоры — терморезистентны благодаря кальциевой соли дипиколиновой кислоты, составляющей 10—15% общей массы споры (в вегетативных клетках эта кислота не встречается). Жизнеспособность спор обеспечивается в течение сотен лет. Попадая в свежую питательную среду, споры прорастают, давая начало новым вегетативным бактериальным клеткам.
Спорообразование (споруляция) начинается сразу после возникновения дефицита питательных веществ в окружающей среде или
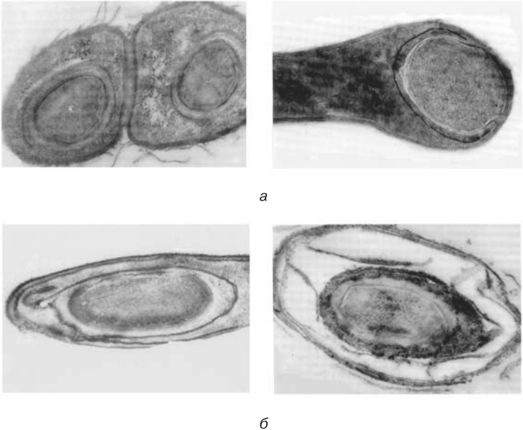
Рис. 3.18. Электронные микрофотографии эндоспор:
а — Bacillus anthracis. Споры формируются в двух еще не разделившихся клетках. Сформировались споровые оболочки и экзоспориум (увел. 80 000 х);
б — Clostridium tetani. Формирование споровой оболочки. Еще хорошо видны элементы цитоплазмы и нуклеоида будущей споры (увел. 95 000 х) (фото В.В. Гузева)
высокой концентрации продуктов обмена веществ старых культур бактерий. Например, у бацилл сибирской язвы споры образуются при температуре не ниже 12 и не выше 42°С и только при доступе кислорода; у бацилл — анаэробов, размножающихся без доступа кислорода воздуха, спорообразование наступает только в отсутствие кислорода. Никаких внешних источников питания или энергии при этом не требуется. Споруляцию стимулирует внесение в среду глюкозы, фосфора и NH4; угнетает внесение пептона, лактозы, NaCl. Споруляция контролируется особыми генами; их число вариабельно у различных видов и может достигать 70. Для спорообразования важна индукция гена spoO.
Процесс спорообразования у разных микроорганизмов происходит с различной скоростью, в среднем для споруляции требуется от 4 до 8 часов.
Академик А.А. Имшенецкий наблюдал у Вас. mycoides спорообразование в течение 70—75 мин. Спорообразование — один из сложнейших процессов дифференцировки бактериальной клетки. Оно начинается с особого неравного деления клетки, увеличения количества белка в определенной части клетки и потери значительного количества свободной воды. Цитоплазма клетки уплотняется и увеличивается количество липидных включений — это подготовительная стадия. Затем бурно начинается стадия пред- споры или образование впячивания цитоплазматической мембраны с измененной плотностью и тинкториальными свойствами (подобное образование называют еще «примордиальная спора»). Третья стадия включает появление оболочки (обычно в течение 10 мин после образования предспоры) и еще большее уплотнение. Стадия созревания споры сопровождается окончательным уплотнением содержимого и снижением метаболической активности клетки. На рис. 3.19 представлен цикл развития спорообразующих бактерий.
Расположение спор (рис. 3.20) может быть центральным, субтерминальным или терминальным. Диаметр споры равен диаметру клетки, в которой она образовалась, — это бациллярный тип спорообразования, характерен для возбудителя сибирской язвы. В ряде случаев клетка приобретает весьма характерный вид: спора формируется на конце клетки, которая при этом несколько расширяется, приобретая вид барабанной палочки, например у возбудителя столбняка плектридиалъный тип спорообразования. У других бактерий спора образуется в центре клетки, которая расширяется в середине, принимая вид веретена. Этот тип называется клостри- диалъным типом спорообразования, например у азотфиксирующих клостридий (С/, pasteurianum).
Прорастанию споры предшествует поглощение ею воды и набухание; происходят глубокие физиологические изменения: дыхание и ферментативная активность быстро возрастают; начинается выделение аминокислот, дипиколиновой кислоты и пептидов (потеря сухой массы споры может достигать 20—30%). Споры теряют свою термоустойчивость. Для перехода споры в вегетативную форму требуется обычно несколько часов (2—3), но иногда достаточно 40—50 мин. Оболочка споры при этом постепенно набухает, увеличивается в размерах, затем разрушается. Чаще всего она разрывается в одном месте, откуда выходит ростовая трубка, из которой затем развивается бактериальная клетка.

Рис. 3.19. Цикл развития спорообразующих бактерий:
В — деление вегетативных клеток; С — свободная зрелая спора;
П — прорастание споры;
- 1—7 — стадии спорообразования: 1 — образование осевого хроматинового тяжа; 2 — образование споровой перегородки (септы); 3 — «поглощение» протопластом материнской клетки септированного участка цитоплазмы с ядром; 4 — формирование кортекса; 5 — начало формирования оболочки споры; 6 — завершение образования оболочки и созревание споры;
- 7 — лизис материнской клетки и освобождение спор(по В.И. Дуде)
Различают полярное и латеральное прорастание споры. При полярном прорастании проросток проходит через отверстие на одном конце и реже — на обоих, при латеральном спора прорастает в средней части клетки, т.е. перпендикулярно к длине оси (рис. 3.21).
Рис. 3.20. Типичные формы спорообразующих клеток:
- 1 — спора расположена в центре, материнская клетка не увеличена(Bacillus megaterium)',2 — спора расположена терминально, материнская клетка не увеличена, заметны белковые включения(Bacillus thuringiensis)
- 3 — спора расположена терминально, материнская клетка раздута в форме булавы(Bacillus polymyxa); 4 — спора расположена в центре, материнская клетка деформировалась и приобрела форму веретена — клостридиальная форма(Bacillus polymyxa); 5 — спора расположена терминально, круглая материнская клетка имеет форму барабанной палочки — плектридиальная форма(Bacillus sphaericus)6 — спора расположена латерально, материнская клетка приобрела веретенообразную форму
- (Bacillus laterosporus)
Рис. 3.21. Прорастание споры по Фурману: а — полярное прорастание; б — латеральное прорастание
Н.А. Красильников допускает, что спорообразование у бактерий является биологической формой обновления организма, способом повышения жизнеспособности клеток и, следовательно, всего вида.
Покоящиеся формы (бактериоцисты, экзоспоры, миксоспоры). Специализированные анабиотические формы известны лишь для ограниченного круга микроорганизмов. Они образуют относительно устойчивые к неблагоприятным условиям клетки — цисты, не являющиеся спорами (например, у азотобактера). Внеклеточные микробные метаболиты, проявляющие свойства индукторов анабиоза, вызывают образование цистоподобных рефрактерных клеток (термин связан с переходом целой клетки в покоящуюся форму и увеличенной способностью данных клеток сильно преломлять свет). В этих клетках отмечается снижение метаболической активности и резистентность к экстремальным воздействиям. Они обладают также рядом особенностей ультраструктурной организации: цитоплазма приобретает мелкозернистую структуру, увеличена толщина клеточной стенки, появляются некоторые плотные включения. Для микроорганизмов, не образующих споры, цистоподобные рефрактерные клетки — единственная покоящаяся форма сохранения вида в течение нескольких лет, а для спорообразующих бактерий — альтернатива спорообразования.
Читайте также: