Дробление яйцеклетки. Этапы дробления яйцеклетки.
Обновлено: 25.04.2024
Основные этапы эмбрионального развития, присущие всем видам животным.
Дробление, типы дробления зародыша у различных позвоночных.
Бластула, виды бластул.
Определение понятия «гаструляция».
Основные способы протекания гаструляции.
Зародышевые листки и их дифференцировка.
I. Электронные микрофотографии
Дробление яйца морского ежа
Дробление яйца лягушки
Ранняя гаструла лягушки
Поздняя гаструла лягушки
II. Таблицы и схемы
Типы дробления яйцеклеток
Дробление яйца ланцетника
Дробление яйца лягушки
Дробление диска яйца курицы
Первые стадии спирального дробления
Гаструляция у лягушки
Типы образования мезодермы
Задания и ориентировочные действия
Изучить характер дробления зародыша в зависимости от типа яйцеклетки
Эл. микрофотографии: дробление яйца морского ежа и лягушки
Рассмотреть, определить тип дробления по схеме, записать
Изучить типы дробления яйцеклеток
Схема типов дробления, таблица с видами дробления (по Флориану)
Зарисовать схемы различных типов дробления, привести примеры животных организмов к каждому типу, закончить заполнение таблицы
Сравнить дробление яйцеклетки различных животных Типа хордовых
Схемы типов дробления яиц ланцетника, лягушки, курицы
Зарисовать схемы различных типов дробления
Изучить строение бластул различных типов
Схема типов бластул
Зарисовать схемы различных типов бластул, привести примеры животных организмов к каждому типу
Изучить образование бластулы в зависимости от характера дробления
Эл. микрофотография бластулы лягушки
Изучить типы гаструляции
Схема типов гаструляции
Зарисовать схемы различных типов гаструляции, привести примеры животных организмов к каждому типу
Изучить образование гаструлы на примере лягушки
Микрофотографии ранней и поздней гаструлы лягушки, схема гаструляции у лягушки
Изучить формирование зародышевых листков
Схема типов образования мезодермы
Зарисовать схемы различных типов образования мезодермы, привести примеры животных организмов к каждому типу
Примерные контрольные вопросы:
Что такое дробление? В чем состоит отличие дробления зародыша от митотического деления клетки?
Какие типы дробления характерны для птиц, млекопитающих? Чем обусловлены различия в типах дробления?
Что такое бластула и какие различают виды бластул?
Какие части различают в зародыше на стадии бластулы?
Что такое гаструла и какие различают типы гаструляции?
Какие зародышевые листки вам известны?
В чем состоит теория зародышевых листков?
Что вы понимаете под нейруляцией?
Лабораторная работа № 3 Развитие ланцетника. Дробление, бластула, гаструляция, закладка осевых органов.
Необходимый исходный уровень знаний
Общая характеристика анамний и амниот.
Особенности эмбриогенеза анамний.
Особенности строения ланцетника, как представителя типа хордовых.
Оплодотворение и дробление ланцетника.
Особенности протекания гаструляции у ланцетника.
Закладка осевых органов у ланцетника.
I. Электронные микрофотографии
Ланцетник (поперечный срез в области глотки)
Яйцеклетка в яичнике лягушки
Дробление яйца лягушки
Ранняя гаструла лягушки
Поздняя гаструла лягушки
II. Таблицы и схемы
Дробление яйца ланцетника
Зародыш ланцетника на стадии бластулы, гаструлы и на стадии обособления эмбриональных зачатков.
Схематический рисунок гаструлы ланцетника.
Икринка лягушки после оплодотворения.
Гаструляция у лягушки
Задания и ориентировочные действия
Изучить характер дробления яйцеклетки ланцетника
Схема дробления яйца ланцетника
Рассмотреть, определить тип дробления по схеме, зарисовать
Изучить строение зародыша ланцетника на разных стадиях развития
Схема зародыша ланцетника на стадии бластулы, гаструлы и обособления эмбр. зачатков
Зарисовать схемы различных этапов развития
Рассмотреть внутреннее строение ланцетника
Эл. микрофотография ланцетника (поперечный срез в области глотки)
Рассмотреть, обратить внимание на расположение органов
Изучить характер дробления яйцеклетки лягушки
Эл. микрофотография яйцеклетки лягушки, схема изменения икринки лягушки после оплодотворения
Дробление яйцеклетки. Этапы дробления яйцеклетки.
Дробление яйцеклетки. Этапы дробления яйцеклетки.
Дробление представляет собой серию митотических делений зиготы с образованием многих дочерних клеток (бластомеров) меньшего размера. Митотические деления зиготы, а в последующем — бластомеров происходят с увеличением числа клеток, но без увеличения их массы, поэтому именуются дроблением.
У человека дробление не имеет принципиальных отличий от такового у других представителей позвоночных, однако протекает гораздо медленнее. Дробление полное, или голобла-стическое (борозды дробления проходят через весь зародыш), неравномерное (в результате дробления образуются дочерние клетки — бластомеры неравной величины) и асинхронное (разные бластомеры дробятся с различной скоростью, поэтому зародыш на отдельных стадиях дробления содержит нечетное число клеток).
Первое деление дробления продолжается в среднем около 30 часов, последующие — более кратковременны (около 20-24 часов). В процессе дробления зародыш перемещается по маточной трубе и на 6-е сутки развития попадает в полость матки.

Бластомеры первой генерации у человека, как и зигота, тотипотентны (каждый бластомер способен развиться в полноценный организм). До стадии 8 бластомеров клетки зародыша формируют рыхлую неоформленную группу, и только после третьего деления устанавливают между собой плотные контакты, образуя компактный клеточный шар из 16 бластомеров, именуемый морулой. Компактизация создает условия для развития наружной клеточной массы и внутренней клеточной массы.
Последняя — это материал будущего тела зародыша (эмбриобласта) и внезародышевых органов. Бластомеры наружной клеточной массы — мелкие и многочисленные (их примерно в 10 раз больше, чем клеток внутренней клеточной массы), являются источником развития трофобласта.
Когда морула попадает в проксимальный отдел маточной трубы и далее — в полость матки, через ее прозрачную зону начинает проникать содержащаяся в маточной трубе и матке жидкость. Происходит кавитация морулы. Сначала жидкость накапливается между клетками и образует небольшие промежутки, которые затем сливаются в единую полость внутри морулы (бластоцель). В образовании жидкости и кавитации участвуют также клетки трофобласта, секретирующие жидкость.
С момента появления полости зародыш именуется бластоцистой. Клетки внутренней клеточной массы бластоцисты локализованы на одном из полюсов и обращены в полость. Клетки наружной клеточной массы уплощаются и, ограничивая полость, формируют оболочку бластоцисты — трофобласт. В период перемещения дробящегося зародыша по маточной трубе большое значение имеет тот факт, что сохраняющаяся прозрачная зона предотвращает прилипание бластоцисты к стенкам трубы и зародыш попадает в полость матки. Здесь он освобождается от прозрачной зоны и начинает имплантироваться (погружаться) в слизистую оболочку матки. Имплантация зародыша протекает параллельно с гаструляцией.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Типы дробления
Тип дробления обычно постоянен и характерен для каждого данного вида животных. Морфология дробления животных различается у разных групп животных. Одним из главных факторов, определяющих эти различия, является количество желтка, содержащегося в яйцеклетке, и распределение его по цитоплазме. При отсутствии или при малых и средних количествах желтка в яйцеклетке, в процессы дробления вовлекается все ее содержимое. В яйцеклетках с большим количеством желтка дробится только небольшая часть, свободная от желтка цитоплазмы. Таким образом, в зависимости от желтка по цитоплазме различают два типа дробления: полное (голобластическое) частичное (меробластическое).
Полное дробление характерно всем яйцеклеткам алецитального, олиголецитального и мезолецитального типа. Погое дробление подразделяется на равномерное и неравномерное. Полное равномерное дробление свойственно яйцеклеткам с малым содержанием желтка (олиголецитальные) и с равномерным его распределением по цитоплазме (гомолецитальные или изолецитальные).
Типичным примером полного равномерного дробления является дробление яйцеклетки ланцетника, впервые изученное А.О. Ковалевским. Такое же дробление наблюдается у голотурий (класс иглокожих). Первая борозда дробления проходит в меридиональном направлении от анимального к вегетативному полюсу, деля зиготу на два бластомера. Вторая борозда также проходит в меридиональном направлении, но в плоскости перпендикулярной плоскости первой борозды, приводя к образовании. 4-х бластомеров. Третья борозда дробления пройдет экваториально, но в плоскости перпендикулярной двум первым плоскостям дробления, деля зародыш на 8 бластомеров. Бластомеры, образующиеся при таком дроблении, имеют приблизительно одинаковые размеры. Дальнейшее чередование борозд дробления в меридиональном и экваториальном направлениях, приводит к расположению бластомеров в виде рядов, соответствующих радиусам шара. Дробление с таким расположением бластомеров называется радиальным. Заканчивается дробление образованием бластулы, которая имеет вид полого шара (Рис. 1).
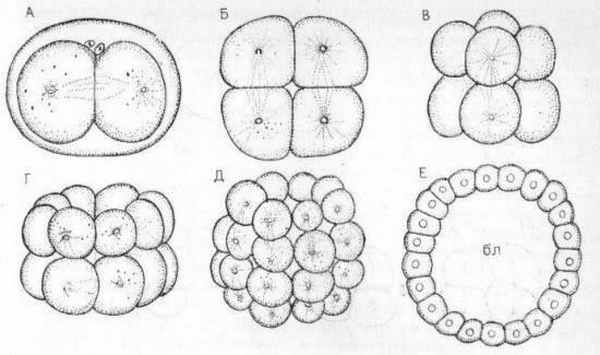
Рис. 1. Последовательные стадии полного равномерного дробления (А-Е) яйца голотурий: Е- целобластула; бл - бластоцель (по Коршельту и Гейдеру).
На первых этапах дробления все бластомеры делятся одновременно (синхронно). При синхронном дроблении число клеток удваивается: 2, 4, 8, 16 и т.д. С течением времени развития синхронность дробления нарушается и становится асинхронным. При асинхронном дроблении темпы деления бластомеров анимального и вегетативного полюсов становятся различными. Продолжительность синхронного и асинхронного периодов у разных животных неодинаковы.
Полное неравномерное дробление характерно яйцеклеткам амфибий, некоторых круглоротых и хрящевых рыб. Рассмотрим этот тип дробления на примере дробления яйцеклетки амфибий. Первые две борозды дробления проходят меридианально, образуя 4-е одинакового размера бластомера. Третья борозда проходит в широтном направлении, ближе к анимальному полюсу и разделяет зародыш на 8 бластомеров не одинаковых размеров. В анимальной части зародыша образуются 4-е мелких бластомера, В дальнейшем происходит чередование меридианальных и широтных борозд дробления, Позднее, у амфибий возникают тангенциальные борозды дробления, деля бластомеры в плоскости, параллельной поверхности зародыша. На заключительном этапе процесса дробления образуется бластула с небольшой полостью, смещенной к анимальному полюсу (Рис. 2.)
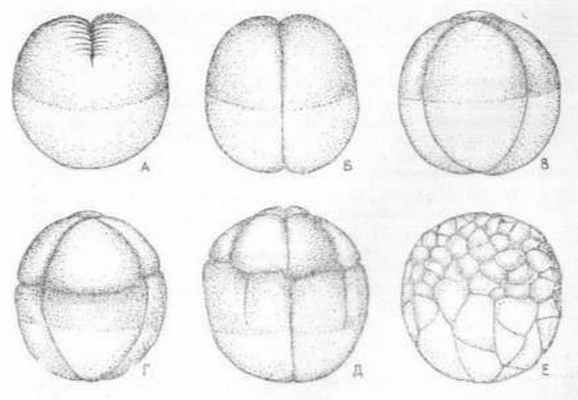
Рис.2. Последовательные стадии дробления (А - Е) яйца лягушки (по Балинскому)
Частичное дробление характерно для полилецитальных яйцеклеток. При частичном дроблении делятся только те части ооплазмы, которые более или менее свободны от желтка, тогда как заполненная часть яйца желтком не подвержена дроблению. В зависимости от распределения желтка в яйцеклетке частичное дробление подразделяется на дискоидальное и поверхностное.
Частичное дискоидальное дробление характерно для телолецитальных яйцеклеток с большим количеством желтка. Такой тип дробления свойственен костистым рыбам, рептилиям и птицам. В процесс дробления вовлекается лишь участок свободной от желтка ооплазмы анимального полюса, где расположено ядро, а другая ее часть, богатая желтком не вовлекается в процесс дробления. Участок активной цитоплазмы яйцеклетки, имеющий форму диска, получил название зародышевого диска. Поэтому и дробление получило название дискоидального. Дискоидальное дробление зародышевого диска начинается с появления меридианальных борозд, которые разделяют зародышевый диск на несколько бластомеров. После этого проходит тангенциальная борозда дробления. Последующие борозды дробления проходят в различных направлениях, и зародышевый диск превращается в многослойную пластинку, которая называется бластодермой. Бластодерма располагается над желтком и отделяется от него узкой щелью - подзародышевой полостью, соответствующей бластоцелю. В результате дробления формируется бластула (Рис. 3).

Рис.3. Последовательные стадии дискоидального дробления (А - Г) яйца курицы. Вид на зародышевый диск сверху (по Белоусову).
Частичное поверхностное дробление характерно центролецитальным яйцеклеткам членистоногих. Дробление центролецитальных яиц происходит таким образом, что после нескольких делений дра, образовавшиеся ядра начинают мигрировать к поверхности яйца. Здесь они располагаются в один ряд, образуя синцитиальный слой. Затем наступает процесс сегментации поверхностного слоя ооплазмы яйцеклетки. В результате поверхностный слой ооплазмы разделяется на бластомеры, образуя бластодерму. Дробление заканчивается образованием бластулы (Рис. 4).
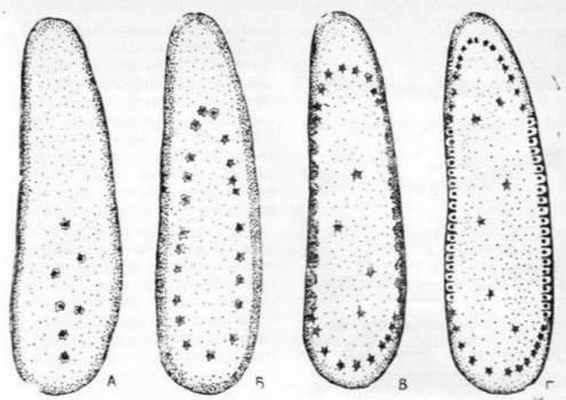
Рис.4. Последовательные стадии (А - Г) поверхностного дробления жука. Ядра дробления постепенно выходят на поверхность яйцеклетки, образуя перибласт (по Белоусову).
Для понимания особенностей разных типов дробления большое значение имеют два правила клеточных делений С. Гертвига и Ю. Сакса.
Согласно первому правилу Гертвига и Сакса ядро яйцеклетки располагается в центре свободной от желтка ооплазмы. Согласно второму праилу, веретено деления ядра располагается в направлении наибольшей протяженности свободной от желтка цитоплазмы. Применимость этих правил для объяснения дробления можно проиллюстрировать на примере дробления яйцеклетки амфибий.
В соответствии с первым правилом Гертвига и Сакса в телолецитальных яйцеклетках амфибий ядро будет располагаться аксцентрично, т.е. смещено к анимальному полюсу яйца. Согласно второму правилу веретено первого деления дробления располагается в широтном направлении (параллельно экватору) яйцеклетки, а борозда дробления в этом случае пройдет перпендикулярно ему. Веретена двух вторых делений яйца будут располагаться в той же плоскости, как и первое, в направлении наибольшей протяженности свободной от желтка цитоплазмы, но под прямым углом к первому веретену. Поэтому в каждом бластомере вторая борозда дробления пройдет перпендикулярно второму веретену деления ядра, т.е. в меридианальном направлении и под прямым углом к плоскости первого дробления. Две меридианальные борозды разделяют зиготу на четыре бластомера. Наибольшая протяженность свободной от желтка цитоплазмы в каждом из них оказывается, теперь в направлении анимально-вегетативной оси и веретена третьих делений дробления будут располагаться в этом направлении (меридианальном), а борозды дробления пройдут в широтной плоскости, перпендикулярно веретанам деления ядра. Последующие борозды дробления будут чередоваться то в меридианальном, то в широтном направлении.
Правила Гертвига и Сакса применимы не только для объяснения процесса дробления яиц амфибий, но и для всех других яйцеклеток телолецитального и центролецитального строения. Однако эти правила не применимы к дроблению алецитальных и олиголецитальных яйцеклеток.
ДРОБЛЕНИЕ
ОСОБЕННОСТИ ДРОБЛЕНИЯ И ОСНОВНЫЕ СТАДИИ ДРОБЛЕНИЯ У МЛЕКОПИТАЮЩИХ
Оплодотворение играет чрезвычайно важную роль в развитии, но оно — лишь только первая его ступень. Зигота с ее новым генетическим потенциалом приступает к созданию многоклеточного организма. У всех животных это начинается с процесса, носящего название дробление — серии митотических делений, в результате которых огромный объем цитоплазмы яйца разделяется на многочисленные клетки меньшего размера. Такие клетки, образующиеся в период дробления, называются бластомерами.
Объем зародыша во время дробления не увеличивается. В большинстве других случаев клеточной пролиферации в период между митозами происходит рост клеток; клетка увеличивается в объеме почти вдвое и затем делится. Такой рост приводит к четкому увеличению общего объема клеток при сохранении относительно постоянного отношения объема ядра к объему цитоплазмы. Однако в период дробления зиготы объем цитоплазмы не возрастает: огромная масса цитоплазмы зиготы разделяется на все более мелкие клетки. Такое деление цитоплазмы яйца, не сопровождающееся ростом, осуществляется путем выпадения интерфазного периода роста между делениями.
Дробление — строго координированный процесс, находящийся под генетическим контролем. Видовые особенности процесса дробления у разных животных определяются двумя основными параметрами: количеством и распределением желтка в цитоплазме и цитоплазматическими факторами, которые влияют на ориентацию митотического веретена.
Наибольшие сложности возникают при изучении дробления у млекопитающих. Яйца млекопитающих относятся к числу самых мелких в животном царстве, что делает затруднительной экспериментальную работу с ними. Диаметр зиготы человека составляет, например, всего лишь 100 мкм, и ее объем меньше одной тысячной объема яйца шпорцевой лягушки. Кроме того, по количеству зигот, которые могут быть получены от одной особи, млекопитающие несравнимы с морскими ежами или лягушками. Обычно у самки млекопитающего одновременно овулирует менее 10 яиц. Кроме того, воспроизведение вне организма матери условий, необходимых для нормального дробления эмбрионов создает дополнительные методические сложности. Однако, к сегодняшнему дню процесс дробления у млекопитающих описан достаточно подробно.
Ооцит млекопитающих, высвобождаясь из яичника, попадает в яйцевод. Оплодотворение происходит в ампуле яйцевода — его отделе, расположенном вблизи яичника. В это время мейоз завершается, и приблизительно через сутки начинается первое деление дробления.
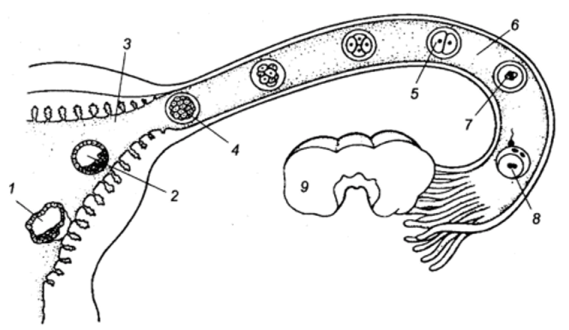
Локализация ранних стадий развития в половых путях самки (Тисктапп Вир1езз в/ а1., 1972; по Гилберт С., 1993):
1 — ранняя стадия имплантации; 2 — бластоциста; 3 — матка; 4 — морула; 5 — двухклеточная стадия; 6 — яйцевод; 7 — первое деление дробления; 8 — оплодотворение; 9 — яичник.
Деления дробления у млекопитающих относятся к числу самых медленных, встречающихся в животном царстве. Каждое из них продолжается от 12 до 24 часов. Тем временем дробящййся зародыш передвигается по яйцеводу по направлению к матке. Первые деления дробления происходят во время перемещения яйца по яйцеводу (рис. 17).
Первая особенность дробления млекопитающих заключается в относительно медленном темпе делений. Второе важное отличие состоит в своеобразном расположении бластомеров относительно друг друга. Первое деление представляет собой нормальное меридиональное деление, т. е. плоскость деления проходит через полюса зиготы. Однако при втором делении один из двух бластомеров делится также меридионально, а второй — экваториально. Такой тип дробления называется чередующимся (рис. 18).
Поскольку яйцеклетки млекопитающих относятся к изо- лецитальному типу, т. е. содержат минимальное количество равномерно распределенного в цитоплазме желтка, для млекопитающих характерно так называемое полное, или голоб- ластическое дробление. Это означает, что борозды дробления проходят через все яйцо. Таким образом, дробление млекопитающих является полным чередующимся асинхронным.

Схема чередующегося дробления млекопитающих (Gulyas, 1975; по Гилберт С., 1993)
Сегодня имеется много данных о том, что компактизация связана с явлениями, происходящими на клеточных поверхностях соседних бластомеров. Во-первых, до компактизации у каждого из восьми бластомеров наблюдаются далеко идущие изменения плазматической мембраны, известные под названием поляризации. Во-вторых, в процессе компактизации участвуют специфические белки клеточной поверхности. Одним таким белком является увоморулин — гликопротеин с молекулярной массой 120 000 дальтон. Антитела к молекуле увоморулина вызывают декомпактизацию морулы и подавляют прикрепление клеток друг к другу. В-третьих, плазматическая мембрана при компактизации может также изменяться вследствие реорганизации цитоскелета. На соседних клеточных поверхностях в результате образования актиновых микрофиламентов возникают микроворсинки, прикрепляющие клетки друг к другу. Именно на ворсинках функционирует увоморулин как медиатор межклеточной адгезии. Уплощение поверхностей соседних бластомеров в месте их контакта может вызываться укорочением микроворсинок путем деполимеризации актина.
Основные стадии доимплантационного развития млекопитающих (по Гилберт С., 1993):
Клетки компактизованного зародыша делятся и образуют 16-клеточную морулу (рис. 19). Такая морула состоит из небольшого числа внутренних клеток, окруженных более многочисленными наружными клетками. Большая часть потомков наружных клеток становится клетками трофобласта (или трофэктодермы). Эта группа клеток не образует эмбриональных структур, а превращается в хорион, участвующий в образовании плаценты. Сам зародыш формируется потомками внутренних клеток 16-клеточного зародыша. Эти клетки образуют внутреннюю клеточную массу (ВКМ), которая и дает начало зародышу. Клетки ВКМ отличаются от клеток трофобласта не только по своему виду, но и по спектру белков, которые они синтезируют на этой ранней стадии. К стадии 64 клеток внутренняя клеточная масса и клетки трофобласта превращаются в полностью сформированные клеточные слои, ни один из которых не поставляет клеток другой группе. Таким образом, возникновение различий между бластомерами трофобласта и внутренней клеточной массы является первым процессом дифференцировки в развитии млекопитающих.
Многочисленные эксперименты показали, что судьба клетки зависит от ее пространственного положения в составе морулы. Если любой бластомер 4-клеточного зародыша мыши поместить на наружную поверхность рыхлой массы бластомеров другого зародыша, то из пересаженной клетки разовьется ткань трофобласта.
Первоначально морула не имеет внутренней полости. Однако в процессе так называемой кавитации клетки трофобласта секретируют в морулу жидкость, что приводит к образованию полости бластоцисты. Внутренняя клеточная масса располагается на одной стороне полого шара, образованного клетками трофобласта. Такая структура называется бластоцистой, и ее образование является еще одной отличительной особенностью дробления млекопитающих.
Пока зародыш перемещается по яйцеводу к матке, бластоциста увеличивается в объеме. В плазматических мембранах клеток трофэктодермы имеется натриевый насос (Ма + /К + -АТФаза), который переносит ионы натрия в центральную полость. Такое накопление ионов натрия вызывает поступление в полость воды осмотическим путем, в результате чего размеры полости бластоцисты увеличиваются. В период перемещения зародыша прозрачная оболочка предотвращает прилипание бластоцисты к стенкам яйцевода. Если у человека происходит такое прилипание, наблюдается «трубная беременность». Это крайне опасное явление, поскольку имплантация зародыша в яйцевод может вызвать угрожающее жизни кровотечение. Однако, когда зародыш достигает матки, он должен освободиться от прозрачной оболочки, чтобы прикрепиться к стенке матки.
Бластоциста мыши освобождается от оболочки путем лизиса в ней небольшого отверстия, через которое она выдавливается при увеличении своего объема. Гистологические данные свидетельствуют о том, что одна из клеток тро- фобластической стенки образует вырост, который приходит в контакт с прозрачной оболочкой. На плазматической мембране этого выроста локализована трипсиноподобная протеаза стрипсин, которая лизирует отверстие в фибриллярном матриксе оболочки. Освободившись от оболочки, бластоциста может вступить в непосредственный контакт с маткой. Здесь трофобласт будет секретировать другую трипсиноподобную протеазу, активатор плазминогена. Этот расщепляющий белок фермент необходим для разрушения ткани матки, чтобы бластоциста могла погрузиться в ее стенку.
1. Онтогенез. Эмбриональное развитие животных
Эмбриональное (зародышевое) развитие охватывает процессы от первого деления зиготы до выхода из яйца или рождения и у большинства животных включает три основных этапа: дробление, гаструляцию и органогенез.
При дроблении дочерние клетки ( бластомеры ) не расходятся и не увеличиваются в размерах. С каждым следующим делением их размеры уменьшаются.
Яйцеклетки с небольшим запасом питательных веществ делятся полностью, т. е. происходит полное дробление. Если яйцеклетка содержит большое количество желтка, то наблюдается частичное дробление — делится только диск цитоплазмы с ядром, а сам желток остаётся без изменений (например, у птиц).
Бластула — это шарообразный зародыш, стенка которого ( бластодерма ) образована одним слоем клеток, а внутри — полость ( бластоцель ).
После дробления начинается гаструляция — часть клеток бластодермы перемещается внутрь зародыша. В результате этих перемещений образуется гаструла.
Гаструла — двухслойный зародыш, состоящий из двух зародышевых листков: наружного ( эктодермы ) и внутреннего ( энтодермы ).
У ланцетника образование гаструлы происходит в результате впячивания части бластодермы внутрь бластоцеля.
Внутренняя полость гаструлы называется первичной кишкой . Её связывает с внешней средой отверстие ( бластопор ), которое становится первичным ртом .

На стадии гаструлы (двух зародышевых листков) прекращается развитие двухслойных животных — губок и кишечнополостных.
У всех остальных животных развитие продолжается, и образуется третий зародышевый листок — мезодерма . Она формируется из энтодермы и всегда расположена между экто- и энтодермой в первичной полости тела.
Дальнейшая специализация клеток зародышевых листков обеспечивает формирование тканей и органов, т. е. гисто- и органогенез .
Из энтодермы образуется хорда — внутренний скелет в виде гибкого тяжа, расположенный на спинной стороне. Позже вместо хорды у позвоночных развивается позвоночник, и только у некоторых животных (например, у хрящевых рыб) её остатки сохраняются в течение всей жизни.
Из эктодермы, расположенной над самой хордой, выделяется нервная пластинка . Затем края пластинки поднимаются и смыкаются. Образуется нервная трубка — зачаток центральной нервной системы. Формируется нейрула .

Нервная трубка, хорда и кишечник создают осевой комплекс органов зародыша, который определяет двустороннюю симметрию тела.

Из эктодермы у позвоночных животных образуется нервная система, органы чувств, покровный эпителий с его железами и производными структурами (волосы, перья, копыта, когти и т. п.).
Из энтодермы формируются органы пищеварительной и дыхательной системы: эпителий средней кишки, печень и поджелудочная железа, жабры, лёгкие, плавательный пузырь, а также щитовидная железа.
Из мезодермы формируются все виды мышечной и соединительной ткани (например, дерма кожи, тела позвонков), кровеносная система, органы выделения, половые железы.
Читайте также: