Рибосомы. Аппарат Гольджи.
Обновлено: 26.07.2024
Мембранные органоиды клетки. Ядро. Прокариоты и эукариоты
Необходимо запомнить
Органоиды клетки
Органоиды, или Органеллы, – постоянные специфические структуры цитоплазмы, выполняющие определённые функции, необходимые для поддержания жизнедеятельности клетки.
Различают органоиды общего значения и специальные органоиды. Органоиды общего значения имеются во всех клетках и выполняют общие функции. Это – митохондрии, рибосомы, эндоплазматическая сеть, комплекс Гольджи, лизосомы, пероксисомы, цитоскелет и клеточный центр.
Органоиды специального значения имеются только в клетках какого-то определённого типа и обеспечивают выполнение функций, присущих только этим клеткам.
Эндоплазматическая сеть (ЭПС) открыта К. Портером в 1945 году. ЭПС или ЭПР (эндоплазматический ретикулум) – сеть канальцев и цистерн, сложенных мембранами. Различают гранулярную (шероховатую, зернистую) и гладкую (агранулярную) ЭПС.

Гранулярная ЭПС содержит рибосомы на наружной стороне мембраны. Гладкая ЭПС не содержит рибосомы. В скелетных мышцах ЭПС носит название саркоплазматический ретикулум. ЭПС пронизывает всю клетку. Полость ЭПС сообщается с ядром и цитоплазматической мембраной.
На рибосомах гранулярной ЭПС синтезируются секреторные белки, предназначенные для выведения из клетки, а также белки лизосом и внеклеточного матрикса.
Наряду с секреторными белками на гранулярной ЭПС синтезируется большая часть полуинтегральных и интегральных белков. В гладеой ЭПС происходит также синтез мембраны липидов и осуществляется «сборка» компонентов мембраны.
Кроме того, ЭПС, как считают, участвует в образовании пероксисом. Таким образом, гранулярная ЭПС служит «фабрикой» мембран для плазмалеммы, аппарата Гольджи, лизосом и других мембранных структур клетки.
Агранулярная (гладкая) эндоплазматическая сеть представляет собой замкнутую сеть трубочек, канальцев, цистерн. На цитоплазматической поверхности гладкой ЭПС синтезируются жирные кислоты, большая часть липидов клетки, в том числе почти все липиды, необходимые для построения клеточных мембран. Поэтому гладкую ЭПС нередко называют «фабрикой липидов». Например, в клетках печени с мембранами гладкого эндоплазматического ретикулума связан фермент, обеспечивающий образование глюкозы из глюкозо-6-фосфата. Эта реакция имеет большое значение в поддержании уровня глюкозы в организме человека.
В организме человека эндоплазматическая сеть особенно хорошо развита в клетках, синтезирующих гормоны, в клетках печени.
Комплекс Гольджи (КГ, или аппарат Гольджи, – пластинчатый комплекс, расположен вблизи ядра, между ЭПС и плазмалеммой. Его структурно-функциональная единица – диктиосома – представляет собой стопку из 5–20 плоских одномембранных мешочков (цистерн), имеющих диаметр около 1 мкм, внутренние полости которых не сообщаются друг с другом. Количество таких мешочков в стопке обычно не превышает 5–20, а расстояние между ними составляет 20–25 нм.

Белки, синтезированные на шероховатой эндоплазматической сети, попадают в аппарат Гольджи. Здесь осуществляется химическая модификация транспортируемых белков и их упаковка в специальные пузырьки.
Таким образом, основными функциями комплекса Гольджи являются химическая модификация, накопление, сортировка, упаковка в секреторные пузырьки и транспорт по назначению белков и липидов, синтезированных в ЭПС.
В комплексе Гольджи образуются лизосомы и синтезируются некоторые полисахариды.
Лизосомальная система и пероксисомы
Лизосомы – мембранные органеллы клеток животных и грибов, содержащие гидролитические ферменты и осуществляющие гидролитическое расщепление макромолекул (внутриклеточное пищеварение). Лизосомы представляют собой окружённые одинарной мембраной пузырьки, размеры которых в клетках животных колеблются от 0,2 до 0,5 мкм. В лизосомах содержится не менее 60 гидролитических ферментов, которые расщепляют все основные классы органических макромолекул.
Все ферменты лизосом активны лишь в кислой среде при значениях pH, близких 5,0. Количество лизосом в разных клетках варьирует от единичных до нескольких сотен, как например, в фагоцитах.
Завершающие этапы процесса внутриклеточного переваривания веществ, поглощённых клеткой, осуществляются в лизосомах.
Лизосомы с помощью своих ферментов могут разрушать не только отдельные органеллы или клетки, но и целые органы (автолиз). Например, в процессе онтогенеза лягушки с помощью ферментов лизосом лизируются хвост и жабры головастика, а образующиеся при этом продукты распада используются для формирования органов взрослого животного.
Митохондрии – крупные мембранные органоиды клетки, которые можно различить в световой микроскоп. Митохондрии присутствуют во всех эукариотических клетках человека, кроме эритроцитов.

Они имеют обычно округлую, удлиненную или нитевидную формы. Количество митохондрий в клетке колеблется в широких пределах (от 1 до 100 тыс. и более) и зависит от потребностей клетки в энергии. Митохондрии имеют наружную и внутреннюю мембраны.
На внутренней поверхности увеличенного фрагмента кристы видны небольшие выпуклости, обращенные в митохондриальный матрикс, которые содержат ферментные системы, обеспечивающие процессы дыхания. Наружная мембрана гладкая и по своему составу сходна с плазмалеммой.
В матриксе содержатся кольцевая молекула митохондриальной ДНК (мтДНК), различные включения, а также молекулы мРНК, транспортной РНК (тРНК) и рибосомы, сходные по строению с рибосомами бактерий. Здесь же располагаются ферменты, превращающие пируват и жирные кислоты в ацетил-КоА, и ферменты реакций цикла Кребса.
Митохондриальная ДНК имеет не линейную, как в хромосомах ядра, а кольцевую форму. Главная функция митохондрий – синтез АТФ, основного источника энергии для обеспечения жизнедеятельности клетки. Поэтому митохондрии называют «энергетическими станциями» клетки.
Пластиды – это органоиды клеток растений и некоторых фотосинтезирующих простейших. У большинства животных и грибов пластид нет.
Пластиды делятся на несколько типов: хлоропласты, хромопласты, лейкопласты. Наиболее важный и известный – хлоропласт, содержащий зелёный пигмент хлорофилл, который обеспечивает процесс фотосинтеза.
Все виды пластид связаны между собой общим происхождением или возможным взаимопревращением. Пластиды развиваются из пропластид – более мелких органоидов меристематических клеток.

Строение пластид
Пластиды относятся к двумембранным органоидам, у них есть внешняя и внутренняя мембраны.
Во многих пластидах, особенно в хлоропластах, хорошо развита внутренняя мембранная система, формирующая такие структуры, как тилакоиды, граны (стопки тилакоидов), ламелы – удлинённые тилакоиды, соединяющие соседние граны. Внутреннее содержимое пластид обычно называют стромой. В ней, помимо прочего, находятся крахмальные зёрна.
Считается, что в процессе эволюции пластиды появились аналогично митохондриям – путём внедрения в клетку-хозяина другой прокариотической клетки, способной в данном случае к фотосинтезу. Поэтому пластиды считают полуавтономными органеллами. Они могут делиться независимо от делений клетки, у них есть собственная ДНК, РНК, рибосомы прокариотического типа, т. е. собственный белоксинтезирующий аппарат. Часть генов, управляющая их функционированием, находится как раз в ядре.
Ядро – важнейшая часть эукариотической клетки. Оно состоит из ядерной оболочки, кариоплазмы, ядрышек, хроматина.
1. Ядерная оболочка по строению аналогична клеточной мембране, содержит поры. Ядерная оболочка защищает генетический аппарат от воздействия веществ цитоплазмы. Осуществляет контроль за транспортом веществ.
2. Кариоплазма представляет собой коллоидный раствор, содержащий белки, углеводы, соли, другие органические и неорганические вещества. В кариоплазме содержатся все нуклеиновые кислоты: практически весь запас ДНК, информационные, транспортные и рибосомальные РНК.
3. Ядрышко – сферическое образование, содержит различные белки, нуклеопротеиды, липопротеиды, фосфопротеиды. Функция ядрышек – синтез зародышей рибосом.
4. Хроматин (хромосомы). В стационарном состоянии (время между делениями) ДНК равномерно распределены в кариоплазме в виде хроматина. При делении хроматин преобразуется в хромосомы.
Функции ядра: в ядре сосредоточена информация о наследственных признаках организма (информационная функция); хромосомы передают признаки организма от родителей к потомкам (функция наследования); ядро согласует и регулирует процессы в клетке (функция регуляции).
Рибосомы. Аппарат Гольджи.
Организация клетки. ЭР и аппарат Гольджи
Устройство и функционирование эндоплазматического ретикулума и аппарата Гольджи

Эндоплазматический ретикулум [ЭР (ER)] — протяженная замкнутая мембранная структура, построенная из сообщающихся трубкообразных полостей и мешочков, называемых цистернами. В области ядра ЭР сообщается с внешней ядерной мембраной. Между шероховатым и гладким ЭР имеется морфологическое различие: мембраны шероховатого ЭР усеяны множеством рибосом, в то время как гладкий ЭР не имеет связанных рибосом.
А. Шероховатый эндоплазматический ретикулум и аппарат Гольджи
Шероховатый ЭР [ШЭР (rER)]( 1 ) — место активного биосинтеза белков. Именно здесь синтезируются белки, которые будут функционировать в составе мембран, лизосом или секретироваться из клетки. Остальные белки синтезируются в цитоплазме на рибосомах, не связанных с мембранами ЭР.
Белки, синтезированные на шероховатом ЭР ( 1 ), претерпевают посттрансляционные модификации (созревание белков, см. с. 226). Они либо остаются внутри шероховатого ЭР в виде мембранных белков, либо транспортируются с помощью везикул ( 2 ) в аппарат Гольджи ( 3 ). Транспортные везикулы образуются почкованием мембран, а затем исчезают, сливаясь с ними (см. с. 230).
Подобно ЭР, аппарат Гольджи ( 3 ) представляет собой сложную сеть ограниченных мембранами полостей, имеющих форму диска и являющихся местом созревания и сортировки белков. Имеются цис -, промежуточная и транс -Гольджи-области и транс -Гольджи-сеть . Посттрансляционная модификация белков имеет место в разных областях аппарата Гольджи.
Наконец, созревшие (модифицированные) белки переносятся везикулами в различные отделы клетки, такие, как лизосомы ( 4 ), цитоплазматическая мембрана ( 6 ) или секреторные пузырьки ( 5 ). Последние высвобождают свое содержимое к межклеточное пространство, сливаясь с плазматической мембраной ( экзоцитоз ). Эти транспортные процессы могут быть конститутивными, т.е. проходить постоянно, или регуляторными, т.е. управляться химическими сигналами. Направленность процесса в первую очередь зависит от сигнальной последовательности синтезируемого белка (см. с. 232).
Наряду с белками в аппарате Гольджи осуществляется транспорт мембранных липидов.
Б. Гладкий эндоплазматический ретикулум
ЭР, не имеющий связанных рибосом, называется гладким эндоплазматическим ретикулумом (ГЭР). Он занимает в клетке Сравнительно небольшой объем. Выраженный ГЭР имеется е клетках с активным обменом липидов, таких, как гепатоциты и клетки Лейдига. Для ГЭР характерна замкнутая система разветвленных канальцев.
ГЭР принимает участие в синтезе липидов . Биосинтез осуществляется ферментами, закрепленными на мембранах ГЭР. Здесь локализован синтез фосфолипидов и отдельные стадии синтеза холестерина (см. с. 174). В ГЭР специализированных клеток эндокринной системы протекают различные стадии синтеза стероидных гормонов (см. с. 364). В ГЭР локализованы также процессы метаболической трансформации ксенобиотиков (реакция 1 , см. с. 308). В этих реакциях принимает участие система цитохрома Р450 (см. с. 310), которую считают основной системой ГЭР.
ГЭР выполняет функцию депо ионов Са 2+ , поддерживающего низкий уровень Са 2+ в цитоплазме. Эта функция более всего свойственна саркоплазматическому ретикулуму, специализированной форме ГЭР мышечных клеток (см. с. 326). В мембранах ГЭР локализованы управляемые Са 2+ -каналы и энергозависимые Са 2+ -насосы, а высокая концентрация ионов Са 2+ в цистернах поддерживается при участии Са 2+ -связывающих белков.
Рибосомы. Аппарат Гольджи.
Рибосомы. Аппарат Гольджи.
Рибосомы — это очень мелкие органеллы (диаметром около 20 нм). Число рибосом в цитоплазме живых клеток весьма велико как у прокариот, так и у эукариот. В обычной бактериальной клетке содержится до 10 000 рибосом, а в эукариотических клетках число их во много раз больше. Рибосомы служат местом синтеза белка.
Каждая рибосома состоит из двух субчастиц, как это можно видеть на рисунке. Из-за мелких размеров рибосомы при дифференциальном центрифугировании седиментируют последними среди всех других органелл: рибосомную фракцию можно получить лишь после центрифугирования при 100 000 g в течение 1—2 ч. Опыты по седиментации выявили существование двух главных типов рибосом, которые были названы 70S- и 80S-рибосомами1. 70S-рибосомы обнаруживаются у прокариот, а несколько более крупные 80S-рибосомы — в цитоплазме эукариотических клеток. Интересно отметить, что в хлоропластах и митохондриях содержатся 70S-pn6ocoMbi, что указывает на какое-то родство этих эукариотических органелл с прокариотами.
Во время синтеза белка на рибосомах аминокислоты последовательно соединяются друг с другом, формируя полипептидную цепь. Подробно этот процесс описан в статьях. Рибосома служит местом связывания для молекул, участвующих в синтезе, т. е. таким местом, где эти молекулы могут занять по отношению друг к другу совершенно определенное положение. В синтезе участвуют: матричная РНК (мРНК), несущая генетические инструкции от клеточного ядра, транспортная РНК (тРНК), доставляющая к рибосоме требуемые аминокислоты, и растущая полипептидная цепь. Кроме того, в этом процессе участвуют факторы инициации, элонгации и терминации цепи. Весь процесс в целом настолько сложен, что без рибосомы он не мог бы идти эффективно (или не шел бы вообще).
В эукариотических клетках отчетливо видны две популяции рибосом — свободные рибосомы и рибосомы, присоединенные к ЭР. Строение тех и других идентично, но часть рибосом связана с ЭР через белки, которые они синтезируют. Такие белки обычно секретируют-ся. Примером белка, синтезируемого свободными рибосомами, может служить гемоглобин, образующийся в молодых эритроцитах.
В процессе синтеза белка рибосома перемещается вдоль нитевидной молекулы мРНК. Процесс идет более эффективно, когда вдоль мРНК перемещается не одна рибосома, а одновременно много рибосом, напоминающих в этом случае бусины на нитке. Такие цепи рибосом называются полирибосомами или полисомами. На ЭР полисомы обнаруживаются в виде характерных завитков.

Аппарат Гольджи
Структуру, известную теперь как аппарат Гольджи, впервые обнаружил в клетках в 1898 г. Камилло Гольджи (Camillo Golgi), применивший в своих наблюдениях особую методику окрашивания. Однако подробно исследовать ее удалось только с помощью электронного микроскопа. Аппарат Гольджи содержится почти во всех эукариотических клетках и представляет собой стопку уплощенных мембранных мешочков, так называемых цистерн, и связанную с ними систему пузырьков, называемых пузырьками Гольджи. Трехмерную структуру аппарата Гольджи трудно выявить при изучении ультратонких срезов, однако предполагают, что вокруг центральной стопки формируется сложная система взаимосвязанных трубочек.
На одном конце стопки постоянно образуются новые цистерны путем слияния пузырьков, отпочковывающихся от агранулярного ЭР. Эта «наружная», или формирующаяся сторона стопки выпуклая, тогда как другая, «внутренняя», где завершается созревание и где цистерны вновь распадаются на пузырьки, имеет вогнутую форму. Стопка состоит из многих цистерн, которые постепенно перемещаются от наружной стороны к внутренней.
Функцию аппарата Гольджи составляют транспорт веществ и химическая модификация поступающих в него клеточных продуктов. Функция эта особенно важна в секреторных клетках, хорошим примером которых могут служить ацинарные клетки поджелудочной железы. Эти клетки секретируют пищеварительные ферменты панкреатического сока в выводной проток железы, по которому они поступают в двенадцатиперстную кишку. На рисунке, А представлена электронная микрофотография такой клетки, а на рисунке схема упомянутого секреторного пути.
Отдельные этапы этого пути выявляют при помощи радиоактивно меченных аминокислот.
Из аминокислот в клетке строятся белки. Используя меченые аминокислоты, можно проследить их включение в белки и передвижение по различным клеточным органеллам. Для этого образцы ткани гомогенизируют через разные промежутки времени после введения аминокислот, разделяют клеточные органеллы центрифугированием и определяют, в каких органеллах наблюдается наивысшая радиоактивность. После концентрирования в аппарате Гольджи белок в пузырьках Гольджи транспортируется к плазматической мембране. Конечным этапом является секреция неактивного фермента посредством процесса, обратного пиноцитозу. Пищеварительные ферменты, выделяемые поджелудочной железой, синтезируются в неактивной форме, чтобы они не могли разрушать клетки, в которых они образуются. Примером может служить трипсиноген, превращающийся в активный трипсин в двенадцатиперстной кишке.
Обычно у белков, поступающих в аппарат Гольджи из ЭР, имеются короткие олигосахаридные цепи, т. е. они представляют собой гликопротеины. Такие углеводные «антенны» могут претерпевать в аппарате Гольджи модификацию, превращающую их в маркеры, с помощью которых белок направляется строго по своему назначению. Однако, каким образом аппарат Гольджи сортирует и распределяет молекулы, в точности не известно.

Иногда аппарат Гольджи участвует в секреции углеводов, например при синтезе материала клеточных стенок у растений. Рисунок свидетельствует об усиленной его активности в области «клеточной пластинки», т. е. в той области, где после деления ядра между двумя только что образовавшимися дочерними ядрами закладывается новая клеточная стенка.
Пузырьки Гольджи направляются к нужному месту на клеточной пластинке при помощи микротрубочек. Мембраны этих пузырьков становятся частью плазматических мембран дочерних клеток, а их содержимое используется для построения срединной пластинки и новых клеточных стенок. Целлюлоза поставляется отдельно, но не через аппарат Гольджи а с помощью микротрубочек.
Два рассмотренных нами примера — секреция ферментов клетками поджелудочной железы и образование новых клеточных стенок в делящихся растительных клетках — показывают, каким образом многие клеточные органеллы могут объединяться для выполнения какой-либо одной функции.
Аппаратом Гольджи секретируется гликопротеин муцин, в растворе образующий слизь. Он выделяется бокаловидными клетками, находящимися в толще эпителия слизистой оболочки кишечника и дыхательных путей. В железах листьев некоторых насекомоядных растений, например росянки, аппарат Гольджи секретиру-ет клейкую слизь и ферменты, с помощью которых эти растения ловят и переваривают добычу. Во многих клетках аппарат Гольджи участвует в секреции слизи, воска, камеди и растительного клея.
Помимо секреции различных веществ аппарат Гольджи выполняет и еще одну важную функцию — в нем формируются лизосомы, к описанию которых мы теперь перейдем.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Лекция № 7. Эукариотическая клетка: строение и функции органоидов
Органоиды — постоянные, обязательно присутствующие, компоненты клетки, выполняющие специфические функции.
Эндоплазматическая сеть
Эндоплазматическая сеть (ЭПС), или эндоплазматический ретикулум (ЭПР), — одномембранный органоид. Представляет собой систему мембран, формирующих «цистерны» и каналы, соединенных друг с другом и ограничивающих единое внутреннее пространство — полости ЭПС. Мембраны с одной стороны связаны с цитоплазматической мембраной, с другой — с наружной ядерной мембраной. Различают два вида ЭПС: 1) шероховатая (гранулярная), содержащая на своей поверхности рибосомы, и 2) гладкая (агранулярная), мембраны которой рибосом не несут.
Функции: 1) транспорт веществ из одной части клетки в другую, 2) разделение цитоплазмы клетки на компартменты ( «отсеки»), 3) синтез углеводов и липидов (гладкая ЭПС), 4) синтез белка (шероховатая ЭПС), 5) место образования аппарата Гольджи.
Аппарат Гольджи
Аппарат Гольджи, или комплекс Гольджи, — одномембранный органоид. Представляет собой стопки уплощенных «цистерн» с расширенными краями. С ними связана система мелких одномембранных пузырьков (пузырьки Гольджи). Каждая стопка обычно состоит из 4-х–6-ти «цистерн», является структурно-функциональной единицей аппарата Гольджи и называется диктиосомой. Число диктиосом в клетке колеблется от одной до нескольких сотен. В растительных клетках диктиосомы обособлены.
Аппарат Гольджи обычно расположен около клеточного ядра (в животных клетках часто вблизи клеточного центра).
Функции аппарата Гольджи: 1) накопление белков, липидов, углеводов, 2) модификация поступивших органических веществ, 3) «упаковка» в мембранные пузырьки белков, липидов, углеводов, 4) секреция белков, липидов, углеводов, 5) синтез углеводов и липидов, 6) место образования лизосом. Секреторная функция является важнейшей, поэтому аппарат Гольджи хорошо развит в секреторных клетках.
Лизосомы
Лизосомы — одномембранные органоиды. Представляют собой мелкие пузырьки (диаметр от 0,2 до 0,8 мкм), содержащие набор гидролитических ферментов. Ферменты синтезируются на шероховатой ЭПС, перемещаются в аппарат Гольджи, где происходит их модификация и упаковка в мембранные пузырьки, которые после отделения от аппарата Гольджи становятся собственно лизосомами. Лизосома может содержать от 20 до 60 различных видов гидролитических ферментов. Расщепление веществ с помощью ферментов называют лизисом.
Различают: 1) первичные лизосомы, 2) вторичные лизосомы. Первичными называются лизосомы, отшнуровавшиеся от аппарата Гольджи. Первичные лизосомы являются фактором, обеспечивающим экзоцитоз ферментов из клетки.
Вторичными называются лизосомы, образовавшиеся в результате слияния первичных лизосом с эндоцитозными вакуолями. В этом случае в них происходит переваривание веществ, поступивших в клетку путем фагоцитоза или пиноцитоза, поэтому их можно назвать пищеварительными вакуолями.
Автофагия — процесс уничтожения ненужных клетке структур. Сначала подлежащая уничтожению структура окружается одинарной мембраной, затем образовавшаяся мембранная капсула сливается с первичной лизосомой, в результате также образуется вторичная лизосома (автофагическая вакуоль), в которой эта структура переваривается. Продукты переваривания усваиваются цитоплазмой клетки, но часть материала так и остается непереваренной. Вторичная лизосома, содержащая этот непереваренный материал, называется остаточным тельцем. Путем экзоцитоза непереваренные частицы удаляются из клетки.
Автолиз — саморазрушение клетки, наступающее вследствие высвобождения содержимого лизосом. В норме автолиз имеет место при метаморфозах (исчезновение хвоста у головастика лягушек), инволюции матки после родов, в очагах омертвления тканей.
Функции лизосом: 1) внутриклеточное переваривание органических веществ, 2) уничтожение ненужных клеточных и неклеточных структур, 3) участие в процессах реорганизации клеток.
Вакуоли
Вакуоли — одномембранные органоиды, представляют собой «емкости», заполненные водными растворами органических и неорганических веществ. В образовании вакуолей принимают участие ЭПС и аппарат Гольджи. Молодые растительные клетки содержат много мелких вакуолей, которые затем по мере роста и дифференцировки клетки сливаются друг с другом и образуют одну большую центральную вакуоль. Центральная вакуоль может занимать до 95% объема зрелой клетки, ядро и органоиды оттесняются при этом к клеточной оболочке. Мембрана, ограничивающая растительную вакуоль, называется тонопластом. Жидкость, заполняющая растительную вакуоль, называется клеточным соком. В состав клеточного сока входят водорастворимые органические и неорганические соли, моносахариды, дисахариды, аминокислоты, конечные или токсические продукты обмена веществ (гликозиды, алкалоиды), некоторые пигменты (антоцианы).
В животных клетках имеются мелкие пищеварительные и автофагические вакуоли, относящиеся к группе вторичных лизосом и содержащие гидролитические ферменты. У одноклеточных животных есть еще сократительные вакуоли, выполняющие функцию осморегуляции и выделения.
Функции вакуоли: 1) накопление и хранение воды, 2) регуляция водно-солевого обмена, 3) поддержание тургорного давления, 4) накопление водорастворимых метаболитов, запасных питательных веществ, 5) окрашивание цветов и плодов и привлечение тем самым опылителей и распространителей семян, 6) см. функции лизосом.
Эндоплазматическая сеть, аппарат Гольджи, лизосомы и вакуоли образуют единую вакуолярную сеть клетки, отдельные элементы которой могут переходить друг в друга.
Митохондрии
Строение митохондрии:
1 — наружная мембрана;
2 — внутренняя мембрана; 3 — матрикс; 4 — криста; 5 — мультиферментная система; 6 — кольцевая ДНК.
Форма, размеры и количество митохондрий чрезвычайно варьируют. По форме митохондрии могут быть палочковидными, округлыми, спиральными, чашевидными, разветвленными. Длина митохондрий колеблется в пределах от 1,5 до 10 мкм, диаметр — от 0,25 до 1,00 мкм. Количество митохондрий в клетке может достигать нескольких тысяч и зависит от метаболической активности клетки.
Митохондрия ограничена двумя мембранами. Наружная мембрана митохондрий (1) гладкая, внутренняя (2) образует многочисленные складки — кристы (4). Кристы увеличивают площадь поверхности внутренней мембраны, на которой размещаются мультиферментные системы (5), участвующие в процессах синтеза молекул АТФ. Внутреннее пространство митохондрий заполнено матриксом (3). В матриксе содержатся кольцевая ДНК (6), специфические иРНК, рибосомы прокариотического типа (70S-типа), ферменты цикла Кребса.
Митохондриальная ДНК не связана с белками («голая»), прикреплена к внутренней мембране митохондрии и несет информацию о строении примерно 30 белков. Для построения митохондрии требуется гораздо больше белков, поэтому информация о большинстве митохондриальных белков содержится в ядерной ДНК, и эти белки синтезируются в цитоплазме клетки. Митохондрии способны автономно размножаться путем деления надвое. Между наружной и внутренней мембранами находится протонный резервуар, где происходит накопление Н + .
Функции митохондрий: 1) синтез АТФ, 2) кислородное расщепление органических веществ.
Согласно одной из гипотез (теория симбиогенеза) митохондрии произошли от древних свободноживущих аэробных прокариотических организмов, которые, случайно проникнув в клетку-хозяина, затем образовали с ней взаимовыгодный симбиотический комплекс. В пользу этой гипотезы свидетельствуют следующие данные. Во-первых, митохондриальная ДНК имеет такие же особенности строения как и ДНК современных бактерий (замкнута в кольцо, не связана с белками). Во-вторых, митохондриальные рибосомы и рибосомы бактерий относятся к одному типу — 70S-типу. В-третьих, механизм деления митохондрий сходен с таковым бактерий. В-четвертых, синтез митохондриальных и бактериальных белков подавляется одинаковыми антибиотиками.
Пластиды

Строение пластид: 1 — наружная мембрана; 2 — внутренняя мембрана; 3 — строма; 4 — тилакоид; 5 — грана; 6 — ламеллы; 7 — зерна крахмала; 8 — липидные капли.
Пластиды характерны только для растительных клеток. Различают три основных типа пластид: лейкопласты — бесцветные пластиды в клетках неокрашенных частей растений, хромопласты — окрашенные пластиды обычно желтого, красного и оранжевого цветов, хлоропласты — зеленые пластиды.
Хлоропласты. В клетках высших растений хлоропласты имеют форму двояковыпуклой линзы. Длина хлоропластов колеблется в пределах от 5 до 10 мкм, диаметр — от 2 до 4 мкм. Хлоропласты ограничены двумя мембранами. Наружная мембрана (1) гладкая, внутренняя (2) имеет сложную складчатую структуру. Наименьшая складка называется тилакоидом (4). Группа тилакоидов, уложенных наподобие стопки монет, называется граной (5). В хлоропласте содержится в среднем 40–60 гран, расположенных в шахматном порядке. Граны связываются друг с другом уплощенными каналами — ламеллами (6). В мембраны тилакоидов встроены фотосинтетические пигменты и ферменты, обеспечивающие синтез АТФ. Главным фотосинтетическим пигментом является хлорофилл, который и обусловливает зеленый цвет хлоропластов.
Внутреннее пространство хлоропластов заполнено стромой (3). В строме имеются кольцевая «голая» ДНК, рибосомы 70S-типа, ферменты цикла Кальвина, зерна крахмала (7). Внутри каждого тилакоида находится протонный резервуар, происходит накопление Н + . Хлоропласты, также как митохондрии, способны к автономному размножению путем деления надвое. Они содержатся в клетках зеленых частей высших растений, особенно много хлоропластов в листьях и зеленых плодах. Хлоропласты низших растений называют хроматофорами.
Функция хлоропластов: фотосинтез. Полагают, что хлоропласты произошли от древних эндосимбиотических цианобактерий (теория симбиогенеза). Основанием для такого предположения является сходство хлоропластов и современных бактерий по ряду признаков (кольцевая, «голая» ДНК, рибосомы 70S-типа, способ размножения).
Лейкопласты. Форма варьирует (шаровидные, округлые, чашевидные и др.). Лейкопласты ограничены двумя мембранами. Наружная мембрана гладкая, внутренняя образует малочисленные тилакоиды. В строме имеются кольцевая «голая» ДНК, рибосомы 70S-типа, ферменты синтеза и гидролиза запасных питательных веществ. Пигменты отсутствуют. Особенно много лейкопластов имеют клетки подземных органов растения (корни, клубни, корневища и др.). Функция лейкопластов: синтез, накопление и хранение запасных питательных веществ. Амилопласты — лейкопласты, которые синтезируют и накапливают крахмал, элайопласты — масла, протеинопласты — белки. В одном и том же лейкопласте могут накапливаться разные вещества.
Хромопласты. Ограничены двумя мембранами. Наружная мембрана гладкая, внутренняя или также гладкая, или образует единичные тилакоиды. В строме имеются кольцевая ДНК и пигменты — каротиноиды, придающие хромопластам желтую, красную или оранжевую окраску. Форма накопления пигментов различная: в виде кристаллов, растворены в липидных каплях (8) и др. Содержатся в клетках зрелых плодов, лепестков, осенних листьев, редко — корнеплодов. Хромопласты считаются конечной стадией развития пластид.
Функция хромопластов: окрашивание цветов и плодов и тем самым привлечение опылителей и распространителей семян.
Все виды пластид могут образовываться из пропластид. Пропластиды — мелкие органоиды, содержащиеся в меристематических тканях. Поскольку пластиды имеют общее происхождение, между ними возможны взаимопревращения. Лейкопласты могут превращаться в хлоропласты (позеленение клубней картофеля на свету), хлоропласты — в хромопласты (пожелтение листьев и покраснение плодов). Превращение хромопластов в лейкопласты или хлоропласты считается невозможным.
Рибосомы
Строение рибосомы:
1 — большая субъединица; 2 — малая субъединица.
Рибосомы — немембранные органоиды, диаметр примерно 20 нм. Рибосомы состоят из двух субъединиц — большой и малой, на которые могут диссоциировать. Химический состав рибосом — белки и рРНК. Молекулы рРНК составляют 50–63% массы рибосомы и образуют ее структурный каркас. Различают два типа рибосом: 1) эукариотические (с константами седиментации целой рибосомы — 80S, малой субъединицы — 40S, большой — 60S) и 2) прокариотические (соответственно 70S, 30S, 50S).
В составе рибосом эукариотического типа 4 молекулы рРНК и около 100 молекул белка, прокариотического типа — 3 молекулы рРНК и около 55 молекул белка. Во время биосинтеза белка рибосомы могут «работать» поодиночке или объединяться в комплексы — полирибосомы (полисомы). В таких комплексах они связаны друг с другом одной молекулой иРНК. Прокариотические клетки имеют рибосомы только 70S-типа. Эукариотические клетки имеют рибосомы как 80S-типа (шероховатые мембраны ЭПС, цитоплазма), так и 70S-типа (митохондрии, хлоропласты).
Субъединицы рибосомы эукариот образуются в ядрышке. Объединение субъединиц в целую рибосому происходит в цитоплазме, как правило, во время биосинтеза белка.
Функция рибосом: сборка полипептидной цепочки (синтез белка).
Цитоскелет
Цитоскелет образован микротрубочками и микрофиламентами. Микротрубочки — цилиндрические неразветвленные структуры. Длина микротрубочек колеблется от 100 мкм до 1 мм, диаметр составляет примерно 24 нм, толщина стенки — 5 нм. Основной химический компонент — белок тубулин. Микротрубочки разрушаются под воздействием колхицина. Микрофиламенты — нити диаметром 5–7 нм, состоят из белка актина. Микротрубочки и микрофиламенты образуют в цитоплазме сложные переплетения. Функции цитоскелета: 1) определение формы клетки, 2) опора для органоидов, 3) образование веретена деления, 4) участие в движениях клетки, 5) организация тока цитоплазмы.
Клеточный центр
Клеточный центр включает в себя две центриоли и центросферу. Центриоль представляет собой цилиндр, стенка которого образована девятью группами из трех слившихся микротрубочек (9 триплетов), соединенных между собой через определенные интервалы поперечными сшивками. Центриоли объединены в пары, где они расположены под прямым углом друг к другу. Перед делением клетки центриоли расходятся к противоположным полюсам, и возле каждой из них возникает дочерняя центриоль. Они формируют веретено деления, способствующее равномерному распределению генетического материала между дочерними клетками. В клетках высших растений (голосеменные, покрытосеменные) клеточный центр центриолей не имеет. Центриоли относятся к самовоспроизводящимся органоидам цитоплазмы, они возникают в результате дупликации уже имеющихся центриолей. Функции: 1) обеспечение расхождения хромосом к полюсам клетки во время митоза или мейоза, 2) центр организации цитоскелета.
Органоиды движения
Присутствуют не во всех клетках. К органоидам движения относятся реснички (инфузории, эпителий дыхательных путей), жгутики (жгутиконосцы, сперматозоиды), ложноножки (корненожки, лейкоциты), миофибриллы (мышечные клетки) и др.
Жгутики и реснички — органоиды нитевидной формы, представляют собой аксонему, ограниченную мембраной. Аксонема — цилиндрическая структура; стенка цилиндра образована девятью парами микротрубочек, в его центре находятся две одиночные микротрубочки. В основании аксонемы находятся базальные тельца, представленные двумя взаимно перпендикулярными центриолями (каждое базальное тельце состоит из девяти триплетов микротрубочек, в его центре микротрубочек нет). Длина жгутика достигает 150 мкм, реснички в несколько раз короче.
Миофибриллы состоят из актиновых и миозиновых миофиламентов, обеспечивающих сокращение мышечных клеток.
Перейти к лекции №6 «Эукариотическая клетка: цитоплазма, клеточная оболочка, строение и функции клеточных мембран»
Строение и функции рибосом, ЭПС, комплекса Гольджи, лизосом
Видеоурок познакомит вас со строением и функциями рибосом, эндоплазматической сети, комплекса Гольджи и лизосом. В уроке рассматривается процесс участия рибосом в синтезе белков, а также понятие сортировки белков в комплексе Гольджи. Основные понятия урока: рибосомы, нуклеотиды, азотистые основания, кодон, антикодон, гранулярная эндоплазматическая сеть, агранулярная эндоплазматическая сеть, автолиз, аутофагия

В данный момент вы не можете посмотреть или раздать видеоурок ученикам
Чтобы получить доступ к этому и другим видеоурокам комплекта, вам нужно добавить его в личный кабинет, приобретя в каталоге.
Получите невероятные возможности
Конспект урока "Строение и функции рибосом, ЭПС, комплекса Гольджи, лизосом"
Рибосомы — это уникальные природные фабрики, синтезирующие белок. Небольшие шарообразные органоиды, диаметр которых составляет 10-30 нм. Рибосомы представляют собой комплексы рибосомальной РНК с белками.

Рибосомы всех живых организмов состоят из двух субъединиц — малой субъединицы, и большой субъединицы.
Рибосомы формируются в ядрышках ядра, а затем выходят в цитоплазму, где и начинают выполнять свою главную функцию – синтез белков.
Рассмотрим процесс участия рибосом в синтезе белков.
Мы ранее говорили о том что генетический код записан на языке нуклеотидов..
В состав которых входят азотистые основания аденин, гуанин, цитозин, которые входят в состав как ДНК, так и РНК. Тимин (T) входит в состав только ДНК, а урацил встречается только в РНК.
Азотистые основания — это 4 строительных кирпичика молекулы ДНК.

Синтез белков начинается с того что в ядре цепь ДНК расплетается с одной из её цепей происходит считывание информации (то есть ДНК выступает как матрица).
Другими словами, происходит перенос генетической информации (ее копирование) с ДНК на РНК.

Такая РНК называется матричной (или информационной) так как она несёт информацию из ядра в цитоплазму на рибосомы.
Сама матричная РНК состоит из кодонов триплетов (в последствии 1 кодон будет кодировать 1 аминокислоту). А из аминокислот как вы знаете состоят белки. И уже в цитоплазме – рибосомы, захватывают матричную РНК.
Однако сперва к матричной РНК присоединяется малая субъединица рибосомы.
Ещё одна важная молекула (транспортная РНК). Она доставляет к рибосомам аминокислоты.
Транспортная РНК выглядит в форме "клеверного листа" и содержит в своём составе тройку нуклеотидов, которую называют антикодоном.
Антикадон взаимосоответствует, то есть комплементарен кодону в матричной РНК с которым он связывается. К концу транспортной РНК присоединена соответствующая аминокислота.
Присоединяясь большая субъединица рибосомы формирует пептидильный (или П-участок) и аминоацильный (или А-участок).
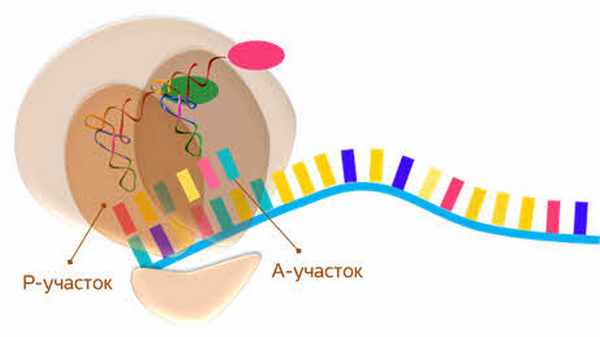
Первая транспортная РНК уходит, и рибосома продвигается дальше до конца матричной РНК.

Таким образом происходит присоединение аминокислот, которые на поверхности рибосомы формируются в полипептидную цепочку.
Таким образом малая субъединица опознает подходящую РНК и место на ней, с которого нужно начать синтез белка. А большая субъединица, содержащая каталитический центр, присоединяется ко всей конструкции и ускоряет образование пептидной связи между растущей полипептидной цепочкой будущего белка и каждой последующей аминокислотой.
Рибосомы могут свободно перемещаться в цитоплазме. Либо прикрепляться к эндоплазматической сети.
Рассмотрим строение и функции эндоплазматической сети.
Эндоплазматическая сеть или эндоплазматический ретикулум как её ещё называют − это внутриклеточный органоид эукариотической клетки, который представляет собой разветвлённую систему из окружённых мембраной уплощённых полостей, пузырьков и канальцев.

Каналы эндоплазматической сети могут пронизывать всю цитоплазму клетки. Они могут ветвиться и соединяться друг с другом, образуя транспортную систему клетки.
Структура эндоплазматической сети не является стабильной и подвержена частым изменениям.
Мембрана эндоплазматической сети морфологически идентична оболочке клеточного ядра и составляет с ней одно целое. Таким образом, её полости открываются в межмембранную полость ядерной оболочки.
Выделяют два вида эндоплазматической сети.
Часть мембран сети покрыта рибосомами – эту часть эндоплазматической сети называют шероховатой (гранулярной).
На шероховатой эндоплазматической сети происходит синтез белков в рибосомах.
Другая часть эндоплазматической сети называется гладкой (агранулярной). Она выполняет транспортную функцию белков.
Посмотрим, как это происходит.
Синтезированный рибосомами белок подходит к мембране эндоплазматической сети в это время она выгибается, захватывает белок и таким образом образуется пузырёк с белком − везикула.
Агранулярная эндоплазматическая сеть также участвует во многих процессах метаболизма. Также она играет важную роль в углеводном обмене, нейтрализации ядов и запасании ионов кальция. Ферменты агранулярной эндоплазматической сети участвуют в синтезе различных липидов и фосфолипидов, жирных кислот и стероидов.
Эндоплазматическая сеть — это транспортная система клетки. В которой происходит синтез белков, липидов и других веществ которые необходимы как самой клетке, так и многим другим клеткам. Если речь идёт о многоклеточно организме.
Также эндоплазматическая сеть принимает участие в том числе и в создании новой ядерной оболочки (например, после митоза).
Строение и функции комплекса Гольджи.
Синтезированные на рибосомах белки проходят через эндоплазматическую сеть и в виде везикул транспортируются в комплекс Гольджи.

Который представлен в виде полостей, уложенных своеобразными стопками.
В отделах комплекса Гольджи белки изменяют свои формы (дозревают). Здесь к ним при помощи специальных ферментов присоединяются такие материалы как липиды и углеводы. Такие белки используются клеткой.
Комплекс Гольджи выполняет роль сортировщика белков. Белки с одинаковой сигнальной последовательностью будут отправлены в одинаковые части клетки, либо за пределы клетки.
Каким же образом это происходит?
От расширений цистерн комплекса Гольджи отщепляются пузырьки, содержащие эти белки. В дальнейшем они могут сливаться друг с другом и увеличиваться в размерах, образуя секреторные гранулы.
После этого секреторные гранулы начинают двигаться к поверхности клетки, соприкасаются с плазмолеммой, с которой сливаются их собственные мембраны, и таким образом содержимое гранул оказывается за пределами клетки.
Так изменённые и сортированные белки выходят из комплекса Гольджи в виде пузырьков.
Из пузырьков комплекса Гольджи содержащей ферменты образуются и (удалить и) пищеварительные органеллы лизосомы.
Рассмотрим строение и функции лизосом.
Лизосома – это окружённый мембраной клеточный органоид, который содержит в себе большой набор ферментов, способных разрушать пищевые вещества.
Данные ферменты формируются из белков в комплексе Гольджи.
Как вы уже знаете клетка захватывает необходимые ей вещества либо при помощи фагоцитоза, либо при помощи пиноцитоза. Поступившие питательные вещества необходимо расщепить (переварить).
Как вы знаете белки расщепляются до аминокислот, полисахариды до глюкозы, а липиды – до глицерина и жирных кислот. И для того что бы, например, полисахарид расщепился, и клетка получила необходимые ей молекулы глюкозы. Ему необходимо встретиться с лизосомой, которая содержит необходимые ферменты для расщепления.
Участие лизосомы в процессе питания простейших, также имеют немаловажную роль. Амёба питается путём фагоцитоза, поглощая бактерий, одноклеточные водоросли и мелких простейших.
Крупные частицы пищи захватываются в пищеварительные вакуоли, которые сливаются с лизосомами. Куда начинают поступать ферменты для переваривания.
Продукты переваривания проникают в цитозоль и используются в качестве пищи. Ферменты лизосомы очень активны, однако они не разрушают окружающую их мембрану.
Иногда лизосомы способны разрушить клетку, в которой они находятся.
Такой процесс разрушения − автолиз просто необходим будущей лягушке.
Лизосомы постепенно переваривают все клетки хвоста головастика во время его превращения в лягушку. А образовавшиеся во время этого процесса вещества всасываются и используются другими клетками тела.

Так же лизосомы можно назвать сборщиками мусора. Они уничтожают повреждённые или изношенные части клетки. К примеру, во время замены старых органоидов новыми, или переваривание белков и других веществ, произведённых внутри самой клетки. Лизосомы своими ферментами расщепляют весь клеточный мусор.
Такой процесс называется аутофагией — уничтожение ненужных клетке структур.
Читайте также: