Поглощение минеральных солей и их транспорт в корне растений.
Обновлено: 26.07.2024
Другой важнейшей функцией корневой системы является поглощение необходимых для растений питательных солей.
Известно, что только молодые растущие части корневой системы (до 5 см от кончика корня) хорошо поглощают минеральные соли. Более старые опробковевшие участки корня не способны к поглощению питательных солей. Таким образом, только при создании хороших условий для роста корня растение нормально обеспечивается питательными веществами.
Корневая система растений -проделывает большую работу, разыскивая и собирая по крупицам рассеянные в почвенной толще питательные элементы. Лишь незначительная часть минеральных солей находится в так называемом почвенном растворе, из которого они без особого труда всасываются корнем. Однако почвенный раствор даже в самой плодородной почве содержит относительно мало питательных веществ и не может обеспечить нормального развития растений. Основная масса питательных солей находится в нерастворимом состоянии или адсорбирована на поверхности мельчайших почвенных частиц.
Питание растений в почве является весьма сложным по сравнению с питанием растений в водных растворах. В почве необходима как большая поверхность корневых систем, так и наличие в ней значительного запаса дыхательных материалов.
Питание растений в почве осложняется еще тем, что питательные вещества, в особенности азот и фосфор, находятся в форме органических соединений. Они становятся доступными для растения лишь после разложения его микроорганизмами, и чем благоприятнее условия для жизнедеятельности микроорганизмов в почве, тем больше питательных веществ получают растения. В холодную погоду разложение органического вещества не идет столь энергично, как летом, поэтому весной растения проявляют зачастую признаки азотного голодания. Вносимый в почву навоз, торф и другие органические удобрения также должны подвергнуться переработке микроорганизмами. Из этого следует, что урожай растений, растущих в почве, до некоторой степени зависит от жизнедеятельности микроорганизмов, К сказанному следует добавить, что зачастую даже плодородные почвы не всегда содержат достаточное количество микроэлементов, которые или отсутствуют или находятся в недоступной для растений форме.
Если рассматривать почву с этих позиций, то каждый почвенный образец представляет собой настолько сложную среду, что часто самый точный химический анализ не % состоянии дать ответ, сколько и каких веществ находится в данный момент в распоряжении растений. Это создает большие трудности гсри решении вопросов, связанных с подкормкой 'При интенсивной культуре растений, так как излишнее внесение минеральных удобрений приводит к засолению почвы и ухудшает ее физические свойства.
Выращивание растений без почвы легко разрешает задачу рационального питания растений, так как в любое время простой количественный анализ позволит дать точный ответ, в каком количестве то или иное вещество находится в растворе. В случае необходимости состав среды можно изменить в нужном направлении. Это является одним из основных преимуществ метода.
Поглощение минеральных веществ является физиологическим процессом и тесно связано с дыханием корней. Поэтому одним из условий интенсивного поглощения питательных солей является хорошая аэрация корневой системы. Очень важны также температурные условия, кислотность и концентрация окружающего раствора. Остановимся отдельно на каждом из этих условий, которые способствуют обеспечению растений питательными веществами.
Смотрите также:
Минеральные вещества почти всегда поглощаются растением в форме ионов.
Типичный опыт на корнях овса показывает, что при повышении концентрации К+ с 0 до 0,02 мМ скорость его поглощения увеличивается линейно, а затем этот прирост.
Из этого следует, что поглощение минеральных веществ клетками корня— процеос избирательный. Кроме того, минеральные вещества обычно накапливаются в тех клетках, где в них возникает потребность.
Поэтому свойства плазмалеммы клеток корня важны для контролирования обмена минеральными веществами между почвой и
Поглощение ионов протекает с использованием энергии АТР и, следовательно, требует аэробных условий в.
Если почвенные частицы упакованы слишком плотно, то объем газовой фазы будет мал, и поглощение минеральных веществ аэробно дышащими корнями снизится из- за недостатка 02.
Корневая система как орган поглощения, усвоения минеральных солей и обмена веществ

Корень — один из основных вегетативных органов листостебельных растений. Ему свойственна хорошо дифференцированная внутренняя структура. Различают стержневую и мочковатую корневую систему. Первая свойственна главным образом двудольным, а вторая — однодольным и некоторым двудольным растениям.
Стержневой корень является непосредственным продолжением стебля. На определенном расстоянии от поверхности почвы у него начинают образовываться боковые, более тонкие корни, которые, в свою очередь, также ветвятся. Мочковатый корень у основания стебля, почти у самой поверхности почвы, образует прядь нитевидных корешков — мочек.
Функции корневой системы многообразны. Она служит для прикрепления растения к субстрату, для поглощения воды и питательных веществ из него, для первичного превращения ряда поглощенных соединений, синтеза органических веществ, которые затем перемещаются в другие органы растения, и для выделения различных продуктов обмена (ионы минеральных солей, аминокислоты, сахара, мочевина, фенольные соединения и др.). Кроме того, у некоторых растений корни выполняют дополнительные функции, например являются вместилищем за-пасных питательных веществ, а у корнеотпрысковых растений — органом вегетативного размножения.
Функции поглощения воды, минеральных веществ и снабжения ими надземных органов у всех растений выполняются одними и теми же структурными элементами корневой системы.
Выросты клеток поверхностной ткани — эпидермиса корня (ризодермис) или эпиблемы поглощающей зоны корня — образуют так называемые корневые волоски. У некоторых растений обнаруживается морфологическая дифференциация эпиблемы на клетки, формирующие волоски (трихобласты), и клетки, которые таких волосков не образуют (атрихобласты). Первые из них короче остальных и отличаются от сестринских клеток, вторые характеризуются более высокой ферментативной активностью и большим содержанием РНК. У растений с гомогенным эпидермисом все клетки эпиблемы обладают способностью к образованию корневых волосков, которые имеют пристенный утолщенный слой цитоплазмы (плазмалемма), ядро и крупную вакуоль.
Тонкая оболочка корневого волоска плотно склеивается с комочками почвы. Корневые волоски служат опорой для растущей верхушки корня, выполняют функцию поглощения воды и минеральных веществ из почвы. Длина корневого волоска 0,06-10 мм, общая же длина корневых волосков, например, растения пшеницы около 20 км. Благодаря корневым волоскам поглощающая поверхность корневой системы возрастает в 5-20 раз. Они выделяют в почву различные вещества, которые способствуют растворению труднодоступных соединений почвы и способствуют развитию микрофлоры. Корневые волоски недолговечны, обычно они отмирают через 15-20 дней.
У многих водных и микотрофных растений, а также в водных культурах корневые волоски отсутствуют; образование их замедляется при ухудшении аэрации и в очень сухой почве. Основная масса корневых волосков сосредоточена в зоне, примыкающей к кончику растущего корня.
Таким образом, функционально поглотительной тканью корня является ризодермис, главным образом корневые волоски, а первичная ассимиляция ионов питательных солей происходит в коре корня.
Между клетками корня могут существовать, значительные электрические градиенты, которые возникают и исчезают, по-видимому, вследствие включения и выключения ионных насосов и изменения концентрационных градиентов в тканях корня.
Исследования показали, что мощность ионных насосов на мембранах корневых волосков значительно выше, чем у соседних клеток эпиблемы без корневых волосков, что обусловливает интенсивное поступление ионов в симпласт.
Первый этап поглощения минеральных солей — пассивное поглощение путем диффузии молекул в клетку и физико-химическая адсорбция их мембраной цитоплазмы — плазмалеммой. На этом этапе минеральные вещества поступают в так называемое «свободное пространство», или «кажущееся свободное пространство», — часть объема клетки или ткани, в которую ионы или молекулы проникают путем свободной диффузии до тех пор, пока их концентрация в «свободном пространстве» не уравновесится с концентрацией внешнего раствора.
Для определения величины свободного пространства (СП) исследуемую ткань погружают на 1 ч в раствор соли известной концентрации для насыщения СП ионами. Затем материал осушают мягкой фильтровальной бумагой и переносят еще на 1 ч в дистиллированную воду для определения экзодиффузии ионов, из СП. Расчет ведут по формуле $$V = \frac100,$$
где V — объем СП, % к общему объему ткани; m — количество ионов, выделившихся за 1 ч на 1 г сырой массы ткани (т. е. примерно из 1 см 3 ), микроэквиваленты; С — концентрация насыщающего раствора, микроэквиваленты/л.
Рассчитанное по калию СП корней ячменя составляет 3-5% общего объема; в молодых тканях на долю СП приходится 15-25% общего объема, что значительно превышает суммарный объем клеточных стенок и межклетников.
Опыты показали, что при перенесении корней из солевого раствора в воду идет экзодиффузия ионов до установления нового равновесия, а при большом количестве воды — до полного, извлечения.
В дальнейшем происходит адсорбция молекул и ионов и активное поступление их в цитоплазму, которое осуществляется: путем обмена ионов Н + и НСО3 - , выделяемых в процессе дыхания цитоплазмой, на адсорбированные анионы и катионы, или перенесение адсорбированных ионов и молекул с помощью, переносчиков в глубоколежащие слои цитоплазмы, где они включаются в метаболизм. При этом анионы поглощаются растениями против градиента электрохимического потенциала, что, требует определенной затраты энергии, а катионы поступают без сопротивления электрических сил.
На перенос поглощенных ионов и молекул оказывает влияние активное движение цитоплазматических мембран клетки. Поглощение крупных органических молекул и капелек растворов может осуществляться с помощью механизма пиноцитоза, при котором на плазматической мембране клетки появляются короткие тонкие выросты, окружающие молекулу или капельку жидкости. Этот участок плазматической мембраны впячивается внутрь протопласта и затем отшнуровывается в виде пузырька. С помощью электронного микроскопа прослежено формирование пиноцитозных пузырьков диаметром 0,07-0,1 мкм.
Д. А. Сабинин, исследуя пасоку, показал, что поглощенные минеральные вещества вступают в химическое взаимодействие с цитоплазмой клеток корня и большая часть их передается в надземные органы в органической форме. Поглощение и передвижение веществ в корне является важным звеном в круговороте элементов в растении и связано с основным звеном обмена — дыханием. Изучение поглощения веществ корневой системой растения дает основание полагать, что единого механизма этого процесса, очевидно, не существует. По-видимому, имеется ряд одновременно функционирующих механизмов поглощения, зависящих от внешних и внутренних факторов.
Установлено, что при водных культурах соли никогда не поступают в корни в той концентрации, в какой они находятся в растворах. Из разбавленных растворов энергичнее поглощаются соли, а из концентрированных — вода. Убедительным доказательством того, что поступление солей не зависит от поглощения воды, служит следующий факт: разные соли и даже анионы и катионы одной и той же соли поступают в растение с различной скоростью. Так, у сульфата аммония (NH4)2SO4 более интенсивно поглощается ион NH4 + , менее интенсивно — ион SO4 2- , в результате чего происходит накопление в растворе ионов SO4 2- и подкисление среды. Поэтому такая соль называется физиологически кислой. Из раствора соли NaNO3 интенсивнее поглощается анион — группа NO3 - , медленнее — катион Na + . В этом случае происходит подщелачивание среды вследствие накопления в ней иона Na + . Такую соль называют физиологически щелочной. У нитрата аммония NH4 + NO3 - с одинаковой интенсивностью поглощаются катионы и анионы. Это физиологически нейтральная соль.
В результате определения электропроводности транспирационного тока воды, поднимающейся по древесине, установлено, что концентрация солей бывает выше ночью, когда транспирация ослаблена и воды в растение поступает меньше. Это также свидетельствует о том, что поступление анионов и катионов соли в живые клетки молодых всасывающих кончиков корней осуществляется независимо от поступления воды. Но при высокой концентрации почвенного раствора транспирационному току принадлежит решающее значение. Он играет большую роль в поднятии воды и минеральных веществ по стеблю.
Поступление веществ в клетку следует рассматривать как активный физиологический процесс, происходящий с затратой определенного количества энергии и связанный прежде всего с дыханием растений. Например, накопление брома в клетках клубней картофеля и в корнях ячменя происходило только тогда, когда через раствор бромида калия, в который были погружены клубни, пропускали в достаточном количестве воздух; при продувании азота через раствор бромида калия поступления и накопления брома в клетках растений не наблюдалось. Добавление сахара к раствору бромида калия усиливало поглощение и накопление калия и брома. Это свидетельствует о том, что поступление ионов связано с процессом дыхания. Считают, что в данном случае главная роль принадлежит ионам Н + и НСО3 - . Угольная кислота, образующаяся в процессе дыхания, направляется в поверхностные слои цитоплазмы, а ионы Н + и НСО3 - обмениваются на катионы и анионы, содержащиеся в окружающей среде. Таким образом, поступление их связано с обменом ионов угольной кислоты на ионы брома и калия.
И. И. Колосов изучал поглощение ионов NO3 - корнями, которые сначала насыщались анионами фосфата. В этом случае поглощение ионов NO3 - сопровождалось освобождением эквивалентного количества H3PO4, ранее адсорбированной на корнях. Об обменной природе адсорбции ионов свидетельствует, например, тот факт, что корни гороха, находящиеся в растворе CаСl2, вместо поглощенного кальция выделяли эквивалентное количество калия и магния.
Основным источником катионов для растений являются обменные катионы почвы. Экзоосмос органических кислот, аминокислот и других соединений корнями — достаточно распространенное явление. Так, в опытах ряда ученых в корневых выделениях кукурузы и других растений были обнаружены сахара (альдозы и кетозы), органические кислоты (яблочная и др.), аминокислоты (аланин, лизин) и витамины (пиридоксин, тиамин, биотин).
Исследованиями В. Ф. Купревича доказано, что высшие растения влияют на субстрат (почву) с помощью ферментов, которые выделяются наружу корневой системой. В свою очередь, наличие корневых выделений высших растений в ризосфере способствует увеличению количества микроорганизмов, обеспечивающих мобилизацию малоподвижных питательных веществ почвы для высших растений. Д. Н. Прянишников установил, что одним из важнейших факторов усвоения аммиачных и нитратных солей является реакция питательной среды. В слабокислой среде при pH 5 нитраты поглощаются более интенсивно, чем аммиачные соли, и, наоборот, в нейтральной среде при pH 7 аммиак поглощается более энергично, чем нитраты.
Отношение растения к аммонийным солям и нитратам зависит от его особенностей. При недостатке углеводов превращение аммонийных солей в амиды задерживается и может вызвать отравление растения аммиаком. При реакции среды, близкой к нейтральной, из соли лучше усваивается катион NH3 + , чем анион NO3 - , и в этом случае соль может из физиологически нейтральной стать физиологически кислой. Ткани лука, погруженные в краску нейтральрот, при pH 4,5 энергично адсорбируют ее оболочками клеток, содержимое которых при этом остается бесцветным. Если перенести клетки в нейтральную среду при pH 7,5, то происходит быстрая десорбция краски оболочкой клеток и переход ее в вакуоль.
Следовательно, поступление и усвоение анионов и катионов минеральных солей корнями в значительной мере зависят от реакции среды.
Усиление кислотности питательного субстрата способствует поглощению анионов. Наоборот, повышение щелочности обусловливает поступление катионов. Так, когда корни поглощают больше анионов, содержание органических кислот в растениях уменьшается, и, наоборот, когда катионы преобладают над анионами, образование органических кислот значительно усиливается. На жизнедеятельность растений pH может оказывать не прямое влияние, а, например, через растворимость в почве питательных веществ. Так, физиологически кислые соли способствуют растворению и усвоению фосфора и фосфоритов, а также солей железа и алюминия. Опытами с водорослями установлено, что чем больше потребность организма в железе, тем в большей мере реакция среды сдвигается в сторону уменьшения pH. Хлороз некоторых растений на щелочных почвах объясняется не вредным действием реакции среды, а недостатком железа в ней. При наличии извести и фосфатов железо переходит в нерастворимую форму и становится недоступным для растений, в результате чего наблюдается хлороз (хотя железо имеется в почве в достаточном количестве); pH почвы может влиять также на растворимость токсического иона алюминия: при pH 4,5-8,5 соли алюминия нерастворимы, а в более кислой среде (pH менее 4,5) растворимость их резко повышается.
Таким образом, поглощение минеральных веществ — физиологический процесс, связанный с общим обменом, поступлением в корневую систему пластических веществ, в частности углеводов, и энергией, а также с особенностями растительного организма и внешними условиями.
Корневую систему сравнительно недавно рассматривали только как орган, функции которого заключаются в поглощении воды и минеральных веществ. Местом синтеза сложных органических веществ считали только лист. И. В. Мичурин одним из первых указал на многогранную роль корней. Он показал, что наследственные свойства привитых растений — качество семян, химический состав, окраска, размеры плодов, форма, опушенность листьев и другие признаки — во многом зависят от природы корней, на которых эти растения развиваются. Корни дикого подвоя всегда ухудшают качество плодов сеянца — привоя и увеличивают процент диких форм в его семенном потомстве. Корни культурного сорта, наоборот, изменяют качество плодов в сторону культурного подвоя. Выводы И. В. Мичурина имели большое практическое значение.
В корнях растений происходят процессы, в результате которых ионы минеральных солей вовлекаются в синтез сложных органических соединений. Это удалось наглядно показать исследованиями пасоки. У кукурузы 50-70% поглощенного корнями азота поступало в надземную часть в виде органических соединений, из них 15-30% приходилось на аминокислоты. В пасоке тыквы обнаружено 16 аминокислот и небольшое количество белка, связанных с солями кальция и кремния. Поглощенный корнями растений СО2 участвует в образовании органических кислот, являющихся акцепторами при синтезе аминокислот.
В опытах А. А. Шмука и Г. С. Ильина с применением 14 С на табаке радиоактивный углерод был найден в никотине лишь у тех растений, которые имели корневую систему. Растения без корней (водные культуры) хотя и поглощали 14 С из раствора NaH 14 CO3, но были неспособны синтезировать молекулы никотина. Выяснено также, что синтез алкалоидов в корнях происходил в том случае, когда растение снабжалось азотом и через листья.
Таким образом, корневая система способна адсорбировать, поглощать и ассимилировать элементы минерального питания, а также передавать продукты своей жизнедеятельности через проводящую сеть в надземные органы.
В растении между корнями и побегами поддерживается постоянный обмен веществами. В некоторых случаях для усиления такого обмена могут образовываться воздушные корни, которые синтезируют аминокислоты (например, у кукурузы). В подземных и воздушных корнях кукурузы происходит синтез аминокислот гистидина, аргинина, аспарагина, серина, глицина, глутаминовой кислоты, аланина и пролина. Корни ее содержат в 10-15 раз больше свободных аминокислот, чем стебли и листья.
В адсорбирующей зоне корней активно идет гликолитический процесс. Под влиянием ферментов сахар превращается в соли пировиноградной кислоты и кислот ди- и трикарбонового цикла, которые являются промежуточными продуктами дыхания корней. Кислоты, переходя друг в друга и постепенно окисляясь обычно до СО2 и воды, создают запас энергии в виде макроэргических фосфатных связей AТФ. Значительная часть этих промежуточных продуктов цикла Кребса используется растением в качестве акцепторов NH2-групп при первичной ассимиляции аммиака. В результате в процессе прямого аминирования и переаминирования кетокислоты превращаются сначала в первичные аминокислоты (аланин, глицин, аспарагиновая и глутаминовая кислоты), а потом частично переходят в более сложные аминокислоты, амиды и другие азотистые соединения. Значительная часть этого материала используется растущими зонами корней для синтеза протеидов и нуклеиновых кислот. Остальные простейшие соединения вместе с пасокой поднимаются из корней в надземные органы, где используются главным образом побегами и развивающимися плодами.
При наличии в ризосфере микроорганизмов в пасоке и корнях обнаруживается большое количество органических соединений азота и фосфора. При этом продукты жизнедеятельности микроорганизмов, выделяющиеся в ризосферу, способствуют не только поглотительной деятельности корневой системы, но и синтетической. Микроорганизмы образуют соединения (витамины и др.), которые могут активизировать рост корней и надземных органов, процессы обмена, дыхания, биосинтез аминокислот и т. д. Бактерии, дрожжи, грибы не только синтезируют необходимые для своей жизнедеятельности витамины, но и выделяют их в окружающую среду. Это одно из звеньев сложных, взаимосвязей между высшими и низшими растениями.
В некоторых почвах обнаружено значительное содержание витаминов. Тиамина особенно много в почвах, в которых находится азотобактер, синтезирующий и в больших количествах выделяющий его в почву. Как выяснилось, в 1 кг навоза содержится 0,13 мг тиамина. Синтез фотохимических пигментов также связан с реакциями, осуществляющимися в корневых системах. Корневая система влияет на активность ферментов, например каталазы и цитохромоксидазы, в листьях. В свою очередь, деятельность корневой системы зависит от надземных органов — листьев, где синтезируются различные пластические и биологически активные вещества.
В жизни растений важная роль принадлежит транспорту ионов. Ионные градиенты, ионные потоки, деятельность ионных насосов в большой мере определяют минеральное питание и обмен веществ. Ионный транспорт важен и для обмена энергией, который происходит с помощью окислительно-восстановительных реакций. В таких реакциях одно вещество (субстрат) окисляется (теряет протоны и электрон), а другое (акцептор) восстанавливается. Энергия, заключенная в химических связях молекул субстрата, освобождается и может быть использована для эндергонических реакций через посредство АТФ.
Наконец, ионный транспорт играет важную роль в информационном обмене. Усиление или ослабление фотосинтеза вызывает соответствующее изменение потребления солей и воды из почвы корневой системой растений. Растение, как известно, обитает в двух средах — корни в почве, а побег в воздухе. Органы, ткани и клетки получают и передают информацию об изменении окружающих условий другим органам, тканям и клеткам. Передача такой информации может осуществляться по симпласту биотоками, возникновение которых связано с распределением катионов и анионов в клетках (Д. Б. Вахмистров, Л. Кларксон).
Поглощение и транспорт минеральных веществ в растении
Корневая система растений поглощает из почвы как воду, так и питательные вещества. Оба процесса взаимосвязаны, но осуществляются на основе разных механизмов. Почти никогда раствор питательных солей не поступает в растение в неизменной концентрации. Из очень разбавленных растворов питательные вещества поглощаются быстрее, чем вода. Наоборот, из концентрированных растворов растение берет больше воды.
Растение поглощает вещества в ионной форме избирательно. Катионы и анионы одной соли поступают в растение относительно независимо друг от друга с разной скоростью. Из солей NH4C1, КС1 больше поглощаются катионы (NH4+, К + ), анионы (СЬ) остаются в растворе и он подкисляется. Эти соли называют физиологически кислыми. Соли NaN03, Са(Н2Р04)2 являются физиологически щелочными. Но можно говорить именно об относительной независимости поглощения. Установлено, что калий лучше усваивается из KN03 чем из K2S04. Фосфат способствует поглощению нитратов.
Установлено, что поглощение веществ происходит не только избирательно, но и против градиента концентраций. Растение накапливает калий, азот, фосфор и другие необходимые для жизнедеятельности элементы в концентрациях, в десятки — сотни раз превосходящих их концентрацию в почвенном растворе.
Процесс поглощения веществ делят на два этапа: поступление ионов в свободное пространство корня и транспорт через мембрану-плазма- лемму.
Свободное пространство корня занимает примерно 10 % его объема и образовано межмолекулярным пространством в толще клеточных стенок.
Клеточные стенки образуют единую систему — апопласт, по которому могут передвигаться растворенные в воде вещества. Кроме того, благодаря наличию заряженных функциональных групп в составе пектиновых веществ, белков и других компонентов клеточной стенки происходит адсорбция и концентрирование ионов. Таким образом, клеточные стенки не только являются фазой транспорта вещества, но и выполняет функцию ионообменников корня. Катионы и анионы поступают в клеточные стенки ризодермы как непосредственно из почвенного раствора, так и благодаря контактному обмену с частицами почвенного поглощающего комплекса (рис. 1.26).
Способность к обменной адсорбции определяется обменной емкостью корня. Она зависит от химического состава корневых выделений и клеточных стенок и поддерживается транспортом веществ через цитоплазматическую мембрану и дальнейшим их перемещением внутрь корня. Транспорт веществ через мембрану может идти по электрохимическому градиенту при участии белков-переносчиков.
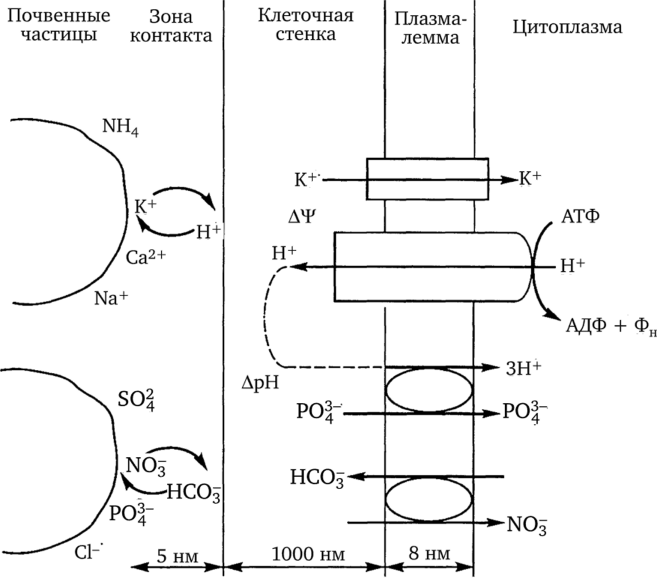
Рис. 1.26. Поглощение веществ клетками ризодермы корня
Особую роль в плазмалемме растительных клеток играет протонная помпа, создающая электрический и химический (pH) градиенты. Электрический потенциал может быть использован для транспорта катионов по электрическому градиенту против концентрационного. В свою очередь, градиент pH служит энергетической основой для переноса через мембрану анионов совместно (в симпорте) с ионами Н+, возвращающимися через мембрану по концентрационному градиенту.
Аналогично Н+ могут функционировать и НС03 или ОН - , избыток которых появляется в примембранном слое цитоплазмы при интенсивной работе Н+-насоса. Транспорт этих ионов и анионов органических кислот наружу по электрохимическому градиенту может протекать в противоположном (антипорте) направлении с поступлением в клетку анионов нитрата.
Именно мембранное строение клеток обеспечивает избирательное поглощение веществ растением. Поддержание структуры мембран и функционирование активных механизмов транспорта ионов являются энергозависимыми процессами. Поэтому существует тесная связь поглотительной деятельности корня с дыханием. Для эффективного функционирования корневой системы она должна обеспечиваться дыхательными субстратами надземной частью. Почвенные условия (аэрация, температура, pH) должны способствовать энергетически эффективному дыханию и росту корней. Установлено, что корневые волоски интенсивно поглощают ионы только до тех пор, пока они растут. Именно наличие большого количества точек роста в корневой системе обеспечивает ее высокую поглотительную активность.
Радиальное перемещение ионов в корне называют ближним транспортом. Он осуществляется через клетки неспециализированных для транспорта веществ тканей по апопласту — совокупности межклетников и межфибриллярных полостей клеточных стенок, симпласту — совокупности протопластов клеток, соединенных плазмодесмами, и вакуому — совокупности дискретных систем вакуолей.
В обычных условиях при сравнительно низких концентрациях ионов в наружной среде поглощенные корневыми волосками ионы поступают в симпласт. Симпластический транспорт ионов по коре сопровождается их включением в процессы обмена веществ. Здесь минеральные формы азота, фосфора и серы включаются в органические соединения и к сосудам уже направляются продукты их первичной ассимиляции. Кора выполняет также функцию транспортного буфера, регулирующего ионный поток корня. При высоком уровне питания (рис. 1.27) избыток ионов сбрасывается в вакуоли и к сосудам подается необходимое количество питательных веществ. При дефицитном питании недостаток ионов смягчается запасами вакуоли.
Особое значение в радиальном транспорте ионов имеет самый внутренний слой клеток коры — эндодерма. Пояски Каспари прерывают апопластический транспорт веществ между корой и центральным цилиндром. Весь поток переходит на симпластический путь, чем обеспечивает метаболический контроль поступления веществ. Следующая ткань — перицикл — работает как кольцевой коллектор, направляя вещества кратчайшим путем из системы симпласта к сосудам ксилемы.
Восходящий транспорт ионов в растении по специализированной проводящей ткани — ксилеме — называют дальним транспортом. Механизм ксилемного транспорта — массовый ток растворенных веществ вместе с водой, обеспечиваемый корневым давлением и транспирацией. Но и на этом этапе транспорта веществ, так же как и в корне, осуществляется его активная регуляция. Применение радиоактивных изотопов фосфора и калия позволило обнаружить, что хотя они и движутся вместе с восходящим транспирационным током, тем не менее в большем количестве поступают не в более энергично транс- пирирующие, а в растущие части растения — в точки роста, только развертывающиеся листочки, в формирующиеся плоды.
Рис. 1.27. Взаимосвязь между симпластным потоком ионов к сосудам ксилемы (горизонтальные стрелки) и ионным обменом между симпластом и вакуумом (вертикальные стрелки) при разных уровнях питания (толщина стрелок условно соответствует интенсивности ионных
БИОЛОГИЯ Том 2 - руководство по общей биологии - 2004
Для питания растениям необходимы не только углеводы, образующиеся при фотосинтезе, но и некоторые минеральные вещества. Для чего нужны эти элементы, указано в табл. 7.7. У высших растений минеральные вещества из почвы или окружающей воды поглощают корни 1 . Максимальное поглощение происходит в зоне корневых волосков. Участие в этом процессе микоризы обсуждается в разд. 7.10.2.
Чтобы понять механизм поглощения и транспорта минеральных ионов, необходимо помнить следующее.
1. Минеральные элементы, необходимые растению, входят в состав солей. В водном растворе молекулы солей диссоциируют и образующиеся ионы свободно передвигаются.
2. Ионы способны пересекать клеточные мембраны различными путями. Один из них — активный транспорт. Он требует затрат энергии в форме АТФ, образующегося в процессе дыхания и может вести к перемещению ионов против градиента их концентрации (разд. 5.9.8).
3. Внутрь корня от его эпидермиса распространяется непрерывная система клеточных стенок — апопласт. Вода и любые растворенные в ней вещества свободно проникают в эту систему из почвы.
На рис. 13.19 приведен график поглощения ионов калия молодыми корнями злаков, предварительно отмытыми чистой водой. Через 90 мин к растворам добавляли ингибитор дыхания — цианид калия.
Рис. 13.19. Поглощение калий-ионов молодым растением злака в аэрируемом растворе.
13.18. а) Опишите, как происходит поглощение ионов калия при 0 и 25 °С.
б) Объясните эффект, вызываемый цианидом калия (KCN).
в) Объясните, зачем надо тщательно отмывать корни перед помещением их в раствор с ионами калия.
Рис. 13.19 показывает, что поглощение ионов четко разделяется на две фазы. Первая длится примерно 10—20 мин. Поглощение в этот период идет относительно быстро. Ионы калия, соприкасаясь с эпидермисом корня, проникают в клеточные стенки и движутся вглубь через апопласт по механизму либо объемного потока, обеспечиваемому транспирацией, либо диффузии. Можно видеть, что эта фаза относительно независима от температуры, поскольку скорость поглощения примерно одинакова и при 25, и при 0 °С. Речь идет о пассивном процессе.
Вторая фаза зависит от температуры и не наблюдается при 0 °С, когда интенсивность метаболизма и дыхания очень низка. Ингибирование поглощения К-ионов цианистым калием свидетельствует о том, что этот процесс зависит именно от дыхания. Во время второй фазы ионы калия поступают в клетки корня через плазма- лемму путем активного транспорта.
Сходные результаты можно получить и на изолированных тканях. Обычно в таких опытах используются запасающие органы растений, например корнеплод моркови. Данные, приведенные на рис. 13.20, подтверждают, что поглощение ионов зависит от дыхания и ингибируется цианидом калия.

Рис. 13.20. Интенсивность дыхания и поглощения хлорида калия дисками из корнеплода моркови. (По данным Robertson, Turner, 1945.)
Итак, поступление ионов в корень обеспечивают два процесса: 1) пассивное поступление, когда ионы движутся за счет объемного потока и диффузии через апопласт; 2) активный транспорт, при котором ионы переносятся в клетки против градиента их концентрации за счет энергии, генерируемой при дыхании.
13.19. На рис. 13.20 видно, что интенсивность дыхания в вырезанных из моркови дисках возрастает, когда их переносят из чистой воды в раствор хлорида калия.
Сравнивая изображенные графики, объясните, почему это происходит.
13.20. Почему поглощение хлорида калия прекращается после добавления KCN?
13.21. Если поставить опыт, аналогичный опыту на рис. 13.19, но с поглощением фосфата, то окажется, что 16% фосфата, поглощенного корнями ячменя, утрачивается в первые несколько минут после перенесения корней в чистую воду. Объясните этот результат.
13.22. Могут ли ионы достичь ксилемы путем передвижения только по апоппасту?
Активный транспорт — процесс избирательный и зависит от дыхания, а диффузия неизбирательна и энергозатрат не требует. В результате пассивного поглощения все клетки первичной коры корня омываются раствором, сходным по составу с почвенным. Так создается обширная поверхность для поглощения ионов.
Ионы, передвигаясь по апопласту, доходят только до эндодермы, где дальнейшее их продвижение блокируют пояски Каспари (разд. 13.5.2). Пересечь этот барьер ионы могут путем диффузии или путем активного транспорта через плазмалеммы эндодермальных клеток и попадая в их цитоплазму и, возможно, в вакуоли. Таким способом растение контролирует, какие минеральные вещества в конце концов попадают в ксилему.
13.23. Каким образом, используя радиоактивные изотопы и радиоавтографию, можно продемонстрировать, что эндодерма является барьером для передвижения ионов через клеточные стенки?
Ионы могут перемещаться и по симпласту. Попав в цитоплазму одной клетки, они будут продвигаться дальше по плазмодесмам, не пересекая мембран. Непрерывный симпластный путь идет от корневых волосков до самой ксилемы. На рис. 13.18, А показаны все возможные пути транспорта ионов внутри корня.
Конечный этап передвижения минеральных солей по корню — высвобождение ионов в ксилему. Для этого им рано или поздно надо покинуть цитоплазму клеток через плазмалемму. Это происходит либо путем диффузии, либо за счет активного транспорта.
Биологическая библиотека - материалы для студентов, учителей, учеников и их родителей.
Наш сайт не претендует на авторство размещенных материалов. Мы только конвертируем в удобный формат материалы, которые находятся в открытом доступе и присланные нашими посетителями.
Если вы являетесь обладателем авторского права на любой размещенный у нас материал и намерены удалить его или получить ссылки на место коммерческого размещения материалов, обратитесь для согласования к администратору сайта.
Разрешается копировать материалы с обязательной гипертекстовой ссылкой на сайт, будьте благодарными мы затратили много усилий чтобы привести информацию в удобный вид.
Поглощение и транспорт корнем воды и минеральных веществ.
Поглощение из почвы воды и доставка к наземным органам – одна из важнейших функций корня, возникшая в связи с выходом на сушу.
Вода попадает в растения через ризодерму, в зоне поглощения, поверхность которой увеличена благодаря наличию корневых волосков. В этой зоне корня формируется ксилема, обеспечивающая восходящий ток воды и минеральных веществ.
Растение поглощает воду и минеральные вещества независимо друг от друга, т.к. эти процессы основаны на различных механизмах действия. Вода проходит в клетки корня пассивно, благодаря осмосу. В корневом волоске находится огромная вакуоль с клеточным соком. Ее осмотический потенциал и обеспечивает поступление воды из почвенного раствора в корневой волосок.
Минеральные вещества поступают в клетки корня в основном в результате активного транспорта. Их поглощению способствует выделение корнем различных органических кислот, переводящих неорганические соединения в доступную для поглощения форму.
В корне горизонтальное движение воды и минеральных веществ происходит в следующей последовательности: корневой волосок, клетки паренхимы коры, эндодерма, перицикл, паренхима осевого цилиндра, сосуды корня. Горизонтальный транспорт воды и минеральных веществ происходит тремя путями:
Путь через апопласт (система, состоящая из межклетников и клеточных стенок). Основной для транспорта воды и ионов неорганических веществ.
Путь через симпласт (система протопластов клеток, соединенная посредством плазмодесм). Осуществляет транспорт минеральных и органических веществ.
Вакуолярный путь – движение из вакуоли в вакуоль через другие компоненты смежных клеток (плазматические мембраны, цитоплазма, тонопласт вакуолей). Применим исключительно для транспорта воды. Для корня незначителен.
В корне вода передвигается по апопласту до эндодермы. Здесь ее дальнейшему продвижению препятствуют пояски Каспари, поэтому дальше вода попадает в стелу по симпласту через пропускные клетки эндодермы. Такое переключение путей обеспечивает регуляцию движения воды и минеральных веществ из почвы в ксилему. В стеле вода не встречает сопротивления и поступает в проводящие сосуды ксилемы.
Вертикальный транспорт воды идет по мертвым клеткам, поэтому перемещение воды обеспечивается деятельностью корня и листьев. Корень подает воду в сосуды стебля под давлением, называемым корневым. Оно возникает в результате того, что осмотическое давление в сосудах корня превышает осмотическое давление почвенного раствора из – за активного выделения клетками корня минеральных и органических веществ в сосуды. Его величина 1 – 3 атм.
Доказательством наличия корневого давления является «плач растения» и гуттация.
«Плач растения» – выделение жидкости из перерезанного стебля.
Гуттация – выделение воды у неповрежденного растения через кончики листьев, когда оно находится во влажной атмосфере или интенсивно поглощает воду и минеральные вещества из почвы.
Верхней силой движения воды является присасывающая сила листьев, обеспечиваемая транспирацией. Транспирация – испарение воды с поверхности листьев. Сосущая сила листьев у деревьев может достигать 15 – 20 атм.
В сосудах ксилемы вода движется в виде непрерывных водяных нитей. Между молекулами воды существуют силы сцепления (когезия), что заставляет их двигаться друг за другом. Прилипание молекул воды к стенкам сосудов (адгезия) обеспечивает восходящий капиллярный ток воды. Основной движущей силой является транспирация.
Для нормального развития растения корни должны быть обеспечены влагой, доступом свежего воздуха и необходимыми минеральными солями. Все это растения получают из почвы, которая представляет собой верхний плодородный слой земли.
Для повышения плодородия почвы в нее вносят различные удобрения. Внесение удобрений во время роста растений называется подкормкой.
Выделяют две основные группы удобрений:
Минеральные удобрения: азотные (селитра, мочевина, сульфат аммония), фосфорные (суперфосфат), калийные (хлорид калия, зола). Полные удобрения содержат азот, фосфор и калий.
Органические удобрения – вещества органического происхождения (навоз, птичий помет, торф, перегной).
Азотные удобрения хорошо растворяются в воде, способствуют росту растений. Их вносят в почву перед посевом. Для созревания плодов, роста корней, луковиц и клубней необходимы фосфорные и калийные удобрения. Фосфорные удобрения плохо растворимы в воде. Их вносят осенью, вместе с навозом. Фосфор и калий повышают холодоустойчивость растений.
Растения в теплицах можно выращивать без почвы, на водной среде, которая содержит все элементы, необходимые растению. Такой способ получил название гидропоники.
Существует также метод аэропоники – воздушной культуры,- когда корневая система находится в воздухе и периодически орошается питательным раствором.
Читайте также: