Рефлекторная регуляция кровообращения.
Обновлено: 02.05.2024
В большинстве структур ЦНС находятся скопления нейронов, стимуляция которых вызывает те или иные изменения функции системы кровообращения. Одна из них представлена в продолговатом мозге, который жестко контролирует деятельность сердечно-сосудистой системы.
Роль продолговатого мозга в регуляции кровообращения В 1871. В. Ф. Овсянников в лаборатории К. Людвига установил, что последовательные фронтальные перерезки продолговатого мозга приводили к существенному снижению артериального давления (до 40-50 мм рт. Ст.). На основании полученных фактов был сделан вывод, что в продолговатом мозге в области дна IV желудочка находятся структуры, ответственные за снижение артериального давления. Дальнейшие исследования установили, что на дорсальной части продолговатого мозга (дно IV желудочка и 3-4 мм ниже) расположены участки, стимуляция которых приводила к незначительному снижению или повышению артериального давления. Эти участки в то время рассматривались как депрессорных и прессорных зоны сосудодвигательного центра, но четкой связи между ними, как по локализации, так и по направленности ответы, установить не удалось.
Только во второй половине XX в. была определена роль и значимость вентральной части продолговатого мозга, как составной части гемодинамического (судинорухово-

РИС. 10.43. Регуляция системного кровообращения
го) центра. На вентральной боковой поверхности ствола мозга, близко к его поверхности, обнаружили скопление нейронов, образовывали три небольших участка - зоны М, S и L, каждая площадью 2 мм2, которые лежат над мягкой мозговой оболочкой и заходят в толщу мозга на 200 600 мкм (рис. 10 44). Они высокочувствительные к химическому (глутамата, глицина, Н2СO3) и электрического раздражения и активно участвуют в регуляции артериального давления и дыхания.
В Ростральные (передний) вентролатерального зоне (РО / 13) ствола мозга (зоны S и М) находятся скопления симпатических нейронов (имеющие выход к спинному мозгу и синтезируют и выделяют адреналин), электрическое или химическое (глютамат) раздражение которых вызывает сужение кровеносных сосудов и повышение артериального давления. После разрушения этих зон или аппликации к ним тормозного медиатора - ГАМК, давление резко и длительно снижается.
В каудальной (задний) вентролатерального зоне (КВЛЗ) (зона L) наблюдаются скопления нейронов (не имеющих выходов к спинному мозгу), электрическое или химическое (глютамат) раздражение которых приводит к снижению артериального давления. их разрушения или аппликация ГАМК вызывают повышение артериального давления.
Таким образом, нейроны ростральной и каудальной вентральной зон продолговатого мозга являются составными частями сосудодвигательного центра. Первые являются симпатоактивуючимы, их возбуждения повышает давление крови вторые, наоборот, снижают артериальное давление путем торможения симпатоактивуючих нейронов.
Рефлекторная регуляция кровообращения К рефлексов, замыкаются в сосудодвигательный центр продолговатого мозга и сопровождаются изменением артериального давления и работы сердца, относятся рефлексы с рефлекторных зон сердечно-сосудистой системы (синокаротидной, аортального, сосудов малого круга кровообращения - легочных артерий и полых вен), имеющих название собственных. Рефлексы, возникающие из других участков тела, называются сопряженными.
Собственные рефлексы возникают с баро- и хеморецепторов рефлексогенных зон вследствие резкого повышения или снижения в них кровяного давления или изменения концентрации химических веществ в крови.
Наиболее исследованным механизмом регуляции артериального давления является барорецепторной рефлексы. Барорецепторы - это рецепторы, раздражение которых происходит при растяжении стенок кровеносных сосудов и сердца. Главную роль в регуляции артериаль-

РИС. 10.44. Структуры сосудодвигательного центра, их локализация и связи. А: 1 - спинной мозг; 2 - продолговатый мозг; 3 - гипоталамус 4 - гипофиз. IX, X - афферентные волокна от барорецепторов каротидного синуса и аорты СН - симпатичная иннервация сосудов и сердца; АВП - аргинин-вазопрессин; CVL - каудальная вентролатерального зона; RVL - ростральная вентролатерального зона; NTS - ядро одинокого тракта (чувствительное ядро блуждающего нерва) ПИЯ - паравентрикулярное ядро; СОЯ - супраоптического ядро, "+" - возбуждающий эффект, - тормозной эффект, ИМЛЯ - интермедиолатеральне ядро спинного мозга, Б; М. S и L - электро- и хемочутливи зоны вентральной поверхности продолговатого мозга ВМ - варолиив городов; X - 10-пара Ч.Н.
ного давления играют барорецепторы каротидного синуса и дуги аорты. Рецепторы синокаротидной зоны содержатся в стенке бифуркации общей сонной артерии на внутреннюю и внешнюю ветви, аорты - в наружном слое стенки ее дуги. Они представлены шишковидной закрученными окончаниями миелиновых нервных волокон. Афферентные волокна от каротидных рецепторов проходят по тонким волокнам нерва Геринга к языкоглоточного нерва и по одинокому тракта - в продолговатый мозг. Максимальная импульсная активность в них возникает при изменении артериального давления в пределах 50-180 мм рт. ст. Менее низкий или более высокий давления крови на изменения импульсной активности не влияют.
Чувствительные волокна от дуги аорты идут по депрессорных нерва (X пара черепных нервов), вливаются в пучок одинокого тракта и заканчиваются в продолговатом мозге. Сенсорные волокна, идущие от синокаротидной зоны и дуги аорты, называют буферными нервами. Сенсорная информация от барорецепторов каротидного синуса поступает как в пресорний, так и в депрессорного центр. От дуги аорты, через сенсорное ядро одинокого тракта, информация поступает только в депрессорного центра, а от хеморецепторов тех же зон - до прессорного центра.
Рефлекторная регуляция кровообращения
4. Симпатическая иннервация сосудов большого круга кровообращения
артерии
Стимуляция симпатических нервов мелких артерий и артериол вызывает
повышение сосудистой сопротивляемости и уменьшению кровотока в
тканях.
Стимуляция симпатических нервов крупных кровеносных сосудов,
особенно вен, приводит к уменьшению объема этих сосудов.
5. Парасимпатическая регуляция
• Играет относительно малую роль.
• Стимуляция парасимпатических нервов
уменьшает частоту сердечных сокращений.
6. Области головного мозга,играющие важную роль в регуляции кровообр-я.
• Сосудодвигательный центр
расположен билатерально в
ретикулярной формации
продолговатого мозга и нижней
трети моста.
• Латеральная частьимпульсы,возбуждающие
симпатические нервы сердца;
• Медиальная частьимпульсы,понижающие частоту и
силу сердечных сокращений
• Сосудодвигательный центр
находится под контролем
вышележащих нервных центров.
ретикулярная
Пунктир-тормозные пути
7. Роль нервной системы в быстрой регуляции АД
Самое главное назначение нервной регуляции
3 механизма:
1. Сужаются практически все артериолы
большого круга кровообращения
2. Происходит значительное сужение вен (и
других крупных сосудов большого круга
кровообращения)
3. Усиление сердечной деят-ти за счет прямого
стимулирующего влияния симпатической н.с.
8. Эффективность нервной регуляции АД
• За 5-10 сек. Давление может
увеличиться в 2 раза по
сравнению со состоянием покоя.
Внезапное торможение нервной
стимуляции сердца и сосудов
,наоборот, может понизить АД на
50% в течение 10-40сек.
9. Рефлекторные механизмы поддержание нормального уровня АД
• Барорецепторный
рефлекс:
• Возникает в ответ на
раздражение рецепторов
растяжения=> их сигнал в
цнс. Затем сигналы
обратной связи
направляются к центрам
автономной нервной
системы и от них к
сосудам. В результате АД
10. Реакция барорецепторов на изменение давления
• Синокаротидные
барорецепторы не
возбуждаются,если Р
имеет величину от 0 до
50-60мм.рт.ст.
• Max частота при Р 180
мм.рт.ст.
• Когда импульсы от
барорецепторов достигают
пучка одиночного тракта в
продолговатом мозге
происходит торможение
сосудосуживающего центра и
возбуждение
парасимпатического центра
блуждающего нерва.
• В результате:
• 1) расширение вен и
артериол в периферических
отделах большого круга
кровоообращения
• 2) уменьшение частоты и
силы сердечных сокращений
12. Хеморецепторные рефлексы
• Эта система реагирует на изменение состава
крови
• Хеморецепторы-клетки,чувствительные к нед.
О2 ,избытку CO2 и ионов водорода.
• Импульсы от хеморецепторов проводятся по
блуждающим нервам и нервам Геринга к
сосудодвигательному центру головного мозга.
• Хеморецепторный рефлекс включается только
после падения давления до 80 мм.рт.ст.
Рефлекторная регуляция кровообращения. Рецепторы ссс. Влияние на гемодинамику коры больших полушарий. Сопряженные рефлексы ссс, их механизмы.
Рефлекторные изменения Рефлекторные изменения сердечной деятельности можно вызвать раздражением рецепторов и других кровеносных сосудов. Например, при повышении давления в легочной артерии замедляется работа сердца. Можно изменить сердечную деятельность и путем раздражения рецепторов сосудов многих внутренних органов.
Обнаружены также рецепторы в самом сердце: эндокарде, миокарде и эпикарде; их раздражение рефлекторно изменяет и работу сердца, и тонус сосудов.
В правом предсердии и у устья полых вен имеются механорецепторы, реагирующие на растяжение (при повышении давления в полости предсердия или в полых венах). Залпы афферентных импульсов от этих рецепторов проходят по центростремительным волокнам блуждающих нервов и вызывают рефлекторное учащение сердечных сокращений. Импульсы, идущие в ЦНС от механорецепторов предсердий, влияют и на работу других органов. Так, при увеличенном наполнении левого предсердия кровью в 2—5 раз возрастает выделение мочи почками, что вызывает уменьшение объема крови и нормализацию наполнения предсердий.
Классический пример вагального рефлекса описал в 60-х годах прошлого века Гольц: легкое поколачивание по желудку и кишечнику лягушки вызывает остановку сердца или замедление его сокращений. Остановка сердца при ударе по передней брюшной стенке наблюдалась также у человека. Центростремительные пути этого рефлекса идут от желудка и кишечника по чревному нерву в спинной мозг и достигают ядер блуждающих нервов в продолговатом мозге. Отсюда начинаются центробежные пути, образованные ветвями блуждающих нервов, идущими к сердцу. К числу вагальных рефлексов относится также глазосердечный рефлекс Ашнера (урежение сердцебиений на 10—20 в минуту при надавливании на глазные яблоки).
Рефлекторное учащение Рефлекторное учащение и усиление сердечной деятельности наблюдается при болевых раздражениях и эмоциональных состояниях: ярости, гневе, радости, а также при мышечной работе. Изменения сердечной деятельности при этом вызываются импульсами, поступающими к сердцу по симпатическим нервам, а также ослаблением тонуса ядер блуждающих нервов.
Условнорефлекторная регуляция сердечной деятельности Тот факт, что различные эмоции вызывают изменение сердечной деятельности, указывает на значение коры полушарий большого мозга в регуляции деятельности сердца. Доказательством этого является то, что изменения ритма и силы сердечных сокращений можно наблюдать у человека при одном упоминании или воспоминании о факторах, вызывающих у него определенные эмоции.
Наиболее убедительные данные о наличии корковой регуляции деятельности сердца получены методом условного рефлекса. Если какой-нибудь, например звуковой, раздражитель сочетать многократно с надавливанием на глазное яблоко, вызывающим уменьшение частоты сердечных сокращений, то затем один этот раздражитель вызывает урежение сердечной деятельности — условный глазосердечный рефлекс.
Условнорефлекторные реакции лежат в основе тех явлений, которые характеризуют так называемое предстартовое состояние спортсменов. Перед соревнованием у них наблюдаются изменения дыхания, обмена веществ, сердечной деятельности такого же характера, как и во время самого соревнования. (У конькобежцев на старте сердечная деятельность учащается на 22—35 сокращений в минуту).
Кора мозга обеспечивает приспособительные реакции организма не только к текущим, но и к будущим событиям. По механизму условных рефлексов сигналы, предвещающие наступление этих событий или значительную вероятность их возникновения, могут вызвать перестройку функций сердца и всей сердечно-сосудистой системы в той мере, в какой это необходимо, чтобы обеспечить предстоящую деятельность организма.
При чрезвычайно сложных ситуациях (действие «чрезвычайных раздражителей», по И. П. Павлову) возможны нарушения и срывы этих корковых высших регуляторных механизмов (неврозы по И. П. Павлову). При этом наряду с расстройствами поведенческих реакций (и невротическими изменениями психологического статуса человека) могут появиться и значительные нарушения деятельности сердца и сердечно-сосудистой системы. В некоторых случаях эти нарушения могут закрепиться по типу патологических условных рефлексов. При этом нарушения сердечной деятельности могут возникнуть при действии одних лишь условных сигналов.
Кафедра нормальной и патологической физиологии с курсом гигиены
1. Физиология как наука, ее задачи и место в системе медицинского образования. История развития физиологии. Методы физиологических исследований. Современные достижения возрастной физиологии. Общие свойства возбудимых тканей. Раздражение и возбуждение. Классификация раздражителей. Закон "силовых отношений" и закон "все или ничего". – 2 часа.
2. Мембранный потенциал и его природа. Структура клеточных мембран. Роль диффузионных и электростатических сил в генезе мембранного потенциала. Электростатические особенности возбудимых тканей в раннем онтогенезе. Потенциал действия, ионный механизм его развития. – 2 часа.
3. Динамика возбудимости при возбуждении. Законы Дюбуа-Реймона: о силе и крутизне нарастания раздражителя, времени его действия. Изменение хронаксии с возрастом. Влияние на ткани постоянного тока. Законы Пфлюгера. – 2 часа.
4. Физиология скелетных мышц. Механизм мышечного сокращения на молекулярном уровне. Функционирование мышц в естественных условиях. Одиночные и тетанические сокращения. Гладкие мышцы, их физиологические особенности.. – 2 часа.
5. Общие свойства центральной нервной системы. Особенности гуморальной, нервной, местной и иммунной регуляции. Нейрон - структурно-функциональная единица нервной системы. Рефлекс, рефлекторная дуга и кольцо. Физиология рецепторов. Основные свойства рецепторов. Механизм развития генераторного и рецепторного потенциалов. Кодирование информации в ЦНС. – 2 часа.
6. Нервный центр. Механизм проведения возбуждения через нервный центр, свойства нервных центров. Возбуждающие медиаторы. – 2 часа.
7. Торможение нервных центров. Тормозные нейроны, их синапсы и медиаторы. Виды и механизмы торможения. – 2 часа.
8. Координация рефлекторной деятельности, ее морфологические и функциональные основы. – 2 часа.
9. Вегетативная (автономная) нервная система. Морфологические и функциональные особенности вегетативной нервной системы. Симпатический, парасимпатический и метасимпатический отделы ВНС, их влияние на функции органов. – 2 часа.
10. Гуморальная регуляция. Общие представления о гуморальном механизме регуляции. Классификация гуморальных агентов и эндокринных желез. Механизмы восприятия и действия гормонов. Контуры саморегуляции, принцип "плюс-минус взаимодействие". Реакция "стресс" и ее значение. – 2 часа.
11. Физиология системы к ровообращения. Сердечный цикл, его фазы. Механические и звуковые проявления сердечной деятельности. – 2 часа.
12. Электрические проявления работы сердца. Особенности биопотенциалов. Электрокардиограмма здорового человека. Генез зубцов, сегментов и интервалов ЭКГ. Значение электрокардиографии в клинике. – 2 часа.
13. Основные законы гемодинамики. Изменение кровяного давления по ходу кровотока. Артериальное давление и способы его измерения. Артериальный пульс, его свойства. Течение крови в венах. – 2 часа.
14. Регуляция работы сердца и сосудистого тонуса. Рефлекторная и гуморальная регуляция кровообращения. – 2 часа.
15. Органное (регионарное) кровообращение и механизмы его регуляции. Изменение системного и органного кровообращения при мышечной нагрузке, приеме пищи, гипоксии, стрессе. Органы – депо крови. Возрастные особенности ССС. – 2 часа.
16. Гистогематический барьер и его значение. Лимфообразование и лимфообращение. Механизм образования тканевой жидкости и собственной лимфы. Регуляция лимфообразования и лимфообращения. – 2 часа.
1. Система дыхания. Внешнее дыхание. Механизм вдоха и выдоха. Легочные объемы и емкости. Функциональные показатели вентиляции легких. Отрицательное давление в плевральной полости и его изменение при дыхании. – 2 часа.
2. Связывание и транспорт О2 и СО2. Кривая диссоциации оксигемоглобина, ее сдвиги влево и вправо. Коэффициент утилизации кислорода. Особенности газообмена и транспорта газов кровью у детей. – 2 часа.
3. Регуляция дыхания. Дыхательный центр, его структура. Гуморальные влияния на дыхательный центр. Собственные и сопряженные рефлексы дыхательной системы. Нереспираторные функции легких. Возрастные особенности регуляции дыхания. – 2 часа.
4. Система в ыделения . Функции почек. Механизм мочеобразования. Регуляция мочеобразования. Антидиуретическая и антинатрийуретическая системы. Искусственная почка. – 2 часа.
5. Физиология системы пищеварения. И.И.Павлов – создатель учения о пищеварении. Функциональная система пищеварения и классификация пищеварительных процессов. Принципы регуляции. Пищевой центр. Иннервация ЖКТ и фазы секреции пищеварительных желез. – 2 часа.
6. Пищеварение в полости рта. Слюнные железы. Состав слюны. Регуляции саливации. Глотание. Пищеварение в желудке. Строение и иннервация желез желудка. Состав желудочного сока. Регуляция желудочной секреции. Особенности пищеварения в полости рта и желудке у детей и в пожилом возрасте. – 2 часа.
7. Пищеварение в двенадцатиперстной кишке. Значение поджелудочной железы и печени. Регуляция панкреатической секреции. Образование и выделение желчи. – 2 часа.
8. Пищеварение в тонком и толстом отделах кишечника. Пристеночное пищеварение /А.М.Уголев/. Роль микрофлоры. – 2 часа.
9. Моторика ЖКТ и ее регуляция. Изменение системы пищеварения в процессе старения. – 2 часа.
10. Обмен веществ и энергии. Обмен веществ и его регуляция. Обмен энергии и методы его изучения. Основной и общий (валовый) обмен энергии. Терморегуляция, ее механизмы: химическая и физическая терморегуляции. Возрастные особенности обмена веществ и энергии. – 2 часа.
11. Частная физиология ЦНС. Физиология спинного и заднего мозга. – 2 часа.
12. Функции среднего и промежуточного мозга. Мозжечок, его функция и последствия его удаления. – 2 часа.
13. Ретикулярная формация. Функции ВРАС и ВРТС, НРАС и НРТС. Тонические рефлексы ствола мозга, их особенности у детей. – 2 часа.
14. Подкорковые ганглии. Кора больших полушарий – высший надсегментарный аппарат ЦНС. Строение коры, представительства функции в коре. Электрические явления в коре. – 2 часа.
15. Современные представления о ноцицептивной и антиноцицептивной системах организма и об их взаимодействии. – 2 часа.
16 Высшая нервная деятельность. /И.П.Павлов, И.М.Сеченов/. Условные и безусловные рефлексы, их особенности и классификации. Механизм образования условных рефлексов. Торможение в коре больших полушарий, виды. Анализ и синтез раздражителей в коре.
17. Современные представления о механизмах памяти. – 2 часа.
18. Осознаваемая и неосознаваемая деятельность мозга. Внимание, виды, характеристика. Физиология сна. Современные представления о механизмах, участвующих в формировании сна и бодрствования. Особенности сна у детей. Физиология гипноза. – 2 часа.
19. Типы ВНД. Особенности ВНД у человека. Первая и вторая сигнальные системы, их взаимоотношения и роль в мышлении. Типологические особенности ВНД детей. Эмоции, происхождение, анатомический субстрат. – 2 часа.
Нервная регуляция кровообращения
Нервная регуляция кровообращения решает несколько важнейших для существования организма задач. Во-первых, она поддерживает такой уровень артериального давления, при котором обеспечивается полноценное кровоснабжение ГМ и сердца. Во-вторых, при условии выполнения первой задачи, снабжает кровью все остальные органы. В-третьих, при условии выполнения двух первых задач, осуществляет перераспределение органного кровотока в соответствии с текущими физиологическими задачами организма (дополняя местные и гуморальные механизмы). Перечисленные задачи решаются с помощью ВНС, способной эффективно влиять на насосную функцию сердца (см. гл. 6) и на тонус сосудов всех органов и тканей.
Нервная регуляция сосудов осуществляется ВНС, главным образом — ее симпатическим отделом. Вегетативные нервы иннервируют все сосуды, кроме капилляров (у них нет гладких мышц — основной мишени нервной регуляции).
Плотность нервных окончаний в разных частях сосудистой системы неодинакова: артерии, как правило, иннервированы лучше, чем вены, а артериолы и мелкие артерии — лучше, чем крупные. Нервные окончания лежат на границе гладкомышечного слоя и наружной оболочки, где они ветвятся, образуя синаптические вздутия (варикозы). Расположение нервных окончаний в стенке артериолы и схема иннервации нреканиллярных сосудов показаны на рис. 8.5.

Рис. 8.5. Схема расположения нервных окончаний в стенке сосуда (а); иннервация микрососудов (б)
Симпатические нервные окончания в большинстве случаев выделяют в синаптическую щель НА, вызывающий сокращение гладкой мускулатуры стенок сосудов. По симпатическим нервным окончаниям к сосудам непрерывно идет поток импульсов с частотой 1—3 в секунду, который вызывает сокращение их гладких мышц. Это сокращение поддерживает сосуды в частично суженом состоянии (нейрогенный сосудистый тонус). Уменьшение импульсации по симпатическим сосудосуживающим нервам приводит к расширению сосудов, усиление имиульсации — к сужению сосудов. Таким образом, наличие нейрогенного тонуса расширяет регуляторные возможности симпатической нервной системы.
У некоторых видов животных, например у собак и кошек, существует особая система симпатических волокон, иннервирующая только артериальные сосуды скелетных мышц. Эта система активируется при стимуляции гипоталамуса. В покое импульсация в этих волокнах отсутствует, но появляется при эмоциональных реакциях: ярости, страхе, тревоге и др. Активация этих волокон приводит к выбросу из нервных окончаний АХ, вызывающего расширение сосудов. Значение этой реакции состоит в подготовке мышц к предстоящей работе. У человека такие волокна пока не обнаружены, хотя расширение сосудов скелетных мышц, предшествующее физической нагрузке, у него наблюдается.
Сосуды наружных половых органов иннервируются волокнами парасимпатического отдела ВНС. В покое импульсация в этих волокнах отсутствует, но появляется при половом возбуждении. Окончания этих волокон выделяют АХ и NO, который расширяет сосуды и увеличивает кровенаполнение половых органов. Парасимпатическую иннервацию имеют также сосуды легких и ГМ.
Таким образом, ведущая роль в регуляции тонуса сосудов принадлежит симпатическому отделу ВНС. Роль парасимпатического отдела незначительна. Это отличает нервную регуляцию сосудов от регуляции сердца, где оба отдела нервной системы одинаково важны.
Преганглионарные симпатические нейроны, участвующие в нервной регуляции сердечно-сосудистой системы, лежат в боковых рогах серого вещества большой группы сегментов спинного мозга (от восьмого шейного до третьего поясничного) и посылают свои аксоны к ганглиям, образующим симпатические цепочки (стволы). В цепочках эти аксоны могут образовывать синапсы на нейронах, лежащих в разных ганглиях. Симпатические ганглионарные нейроны непосредственно иннервируют сердце и сосуды, причем в стенке одного сосуда обычно располагаются нервные окончания, связанные с нейронами из нескольких ганглиев. Основные пути эффек- торного звена нервной регуляции сердечно-сосудистой системы показаны на рис. 8.6.
Активность преганглионариых симпатических нейронов находится под контролем нейронной сети, расположенной в продолговатом мозге и варолиевом мосте. Эта сеть получила название сердечно-сосудистого центра.
Сердечно-сосудистый центр ГМ располагается в ретикулярной формации продолговатого мозга и нижней трети моста. В его состав входит ядро блуждающего нерва и зоны, активация которых вызывает сужение сосудов с одновременным усилением работы сердца (прессорный центр) и расширение сосудов с одновременным угнетением работы сердца (депрес- сорный центр). Нейроны прессорного центра активируют симпатические преганглионарные нейроны и тормозят парасимпатические нейроны ядра блуждающего нерва. Нейроны депрессорного центра тормозят активность нейронов прессорного центра и активируют нейроны ядра блуждающего нерва.

Рис. 8.6. Схема анатомического строения основных путей и центров симпатической регуляции сердечно-сосудистой системы:
пунктиром показан парасимпатический путь в составе блуждающего нерва
Сердечно-сосудистый центр, с одной стороны, получает и интегрирует информацию от многочисленных рецепторов сердечно-сосудистой системы, а с другой — находится под постоянным контролем вышележащих нервных центров (рис. 8.7).
На сердечно-сосудистый центр оказывают влияние рецепторы, измеряющие системное давление крови (барорецепторы); рецепторы, измеряющие уровень кровоснабжения тканей (хеморецепторы 02, С02 и др.); рецепторы, измеряющие состояние стенок сердца (механорецепторы сердца), и др. Сердечно-сосудистый центр взаимодействует с дыхательным центром, который также располагается в структурах продолговатого мозга и моста.
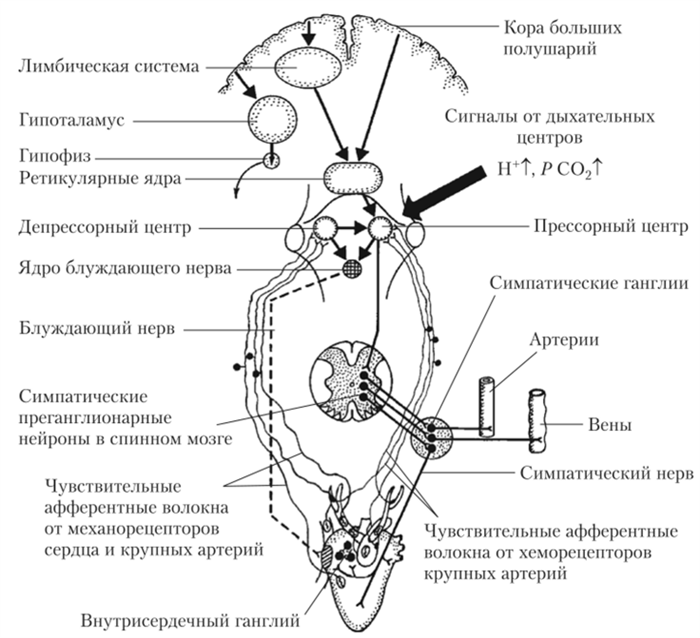
Рис. 8.7. Схема основных связей сердечно-сосудистого регуляторного центра продолговатого мозга
Из вышележащих отделов ГМ на сердечно-сосудистый центр наибольшее влияние оказывают ядра ретикулярной формации моста, среднего мозга и гипоталамуса. На сердечно-сосудистый центр влияют также лимбическая система и некоторые отделы коры больших полушарий: двигательная область, передняя часть височной доли, орбитальная область лобной коры, передняя часть поясной извилины и др. Области ГМ, влияющие на сердечно-сосудистый центр, показаны на рис. 8.8.
Большинство из перечисленных структур могут оказывать на работу сердечно-сосудистого центра как угнетающее, так и стимулирующее влияние в зависимости от интенсивности возбуждения и исходной активности центра.
Барорецепторы, осуществляющие контроль за уровнем давления, располагаются в основном в крупных сосудах и в сердце. Там они образуют скопления, которые называются рефлексогенными зонами. Важнейшими из них являются зоны дуги аорты и каротидного синуса (место ветвления сонной артерии на внутреннюю и внешнюю). Расположение этих зон показано на рис. 8.9.

Рис. 8.8. Области головного мозга, участвующие в регуляции кровообращения, и связывающие их пути

Рис. 8.9. Расположение барорецепторов (БР) и хеморецепторов (ХР) в аорте и сонной артерии
Барорецепторы по сути являются рецепторами растяжения, т.е. механорецепторами, реагирующими на изменения напряжения в стенке сосуда. Барорецепторы активны уже при нормальном уровне давления крови. Снижен не давления во время диастолы желудочков уменьшает их активность, а подъем давления во время систолы желудочков — увеличивает. При возрастании давления выше нормы частота импульсации от барорецепторов прогрессивно возрастает. Сигналы от барорецепторов поступают в депрес- сорный центр продолговатого мозга, который в ответ тормозит активность прессорного центра и активирует блуждающий нерв. В результате сосуды расширяются, а сердце тормозит свою работ>'. В совокупности это приводит к очень быстрому и эффективному снижению давления. Активность барорецепторов при этом тоже снижается. Напротив, при снижении артериального давления частота импульсов, идущих от барорецепторов, уменьшается, что приводит к рефлекторному усилению работы сердца и сужению сосудов. В результате артериальное давление повышается. Таким образом, барорефлексы стабилизируют давление на определенном уровне. Этот уровень задается сердечно-сосудистым центром под влиянием сигналов от других рецепторов или вышележащих отделов ГМ.
Сердечные механорецепторы были описаны в гл. 6.
Артериальные хеморецепторы, расположенные в каротидных и аортальных тельцах (см. рис. 8.9), не только играют важную роль в рефлекторной регуляции дыхания, но и влияют на систему кровообращения.
Хеморецепторы представляют собой клетки, чувствительные к недостатку кислорода, а также избытку углекислого газа и ионов водорода. Эти клетки располагаются в небольших (диаметром около 2 мм) хеморе- цептивных органах. Два из них лежат в области каротидного синуса (каротидные тельца), а два-три — вблизи аорты (аортальные тельца). Импульсы от этих хеморецепторов поступают в сердечно-сосудистый центр по чувствительным нервным волокнам, идущим параллельно с волокнами от барорецепторов (см. рис. 8.9). Хеморецепторы находятся в постоянном контакте с кровью, так как густо оплетены капиллярами. Когда артериальное давление падает, хеморецепторы начинают ощущать недостаток кислорода и избыток углекислого газа и ионов водорода, в результате чего они возбуждаются. Усиленная импульсация от рецепторов активирует сердечно-сосудистый центр, который возвращает давление к исходному уровню.
Таким образом, комплекс баро- и хеморецепторов осуществляет постоянный контроль за уровнем артериального давления, посылая сигналы о его отклонениях от нормы в сердечно-сосудистый центр. Этот центр, оказывая влияние на работу сердца и тонус сосудов, обеспечивает рефлекторную стабилизацию давления. Нейрогенные механизмы при необходимости дополняются гуморальными, в частности — эндокринными, которые также способны участвовать в регуляции работы сердца и тонуса сосуда. Время срабатывания нейрогенных механизмов регуляции — секунды, гуморальных — минуты и часы, что позволяет отнести их к системе быстрой регуляции кровообращения. В организме, однако, существует мощный механизм регуляции артериального давления, эффективность которого проявляется не так быстро, но поддерживается в течение недель и даже месяцев. Этот механизм может быть отнесен к системе долговременной регуляции кровообращения.
Читайте также: