Анатомия и физиология дорсального кохлеарного ядра (ДКЯ)
Обновлено: 02.05.2024
У волосковых клеток нет аксона, они соединены с отростками второго нейрона (биполярной клетки), тело которого лежит в улитке. Слуховые волокна обладают спонтанной активностью (в отсутствие стимуляции). К волосковым клеткам также идут эфферентные волокна от клеток ядра оливы. Если их стимулировать, то это приводит к угнетению ответа воло- сковых клеток.
Слуховой тракт начинается от первичных чувствительных нейронов, которые располагаются в спиральном ганглии вблизи улитки. Аксоны этих нейронов следуют к кохлеарным ядрам среднего мозга, где осуществляется первое синаптическое переключение. Аксоны нейронов этих ядер направляются к ипси- и контралатеральным оливарным комплексам (продолговатый мозг), что обеспечивает интегрирование сигналов от левого и правого уха на этом уровне (меньшая часть волокон остается в своем полушарии, а большая часть переходит на противоположную сторону). Далее, после синаптического переключения в ядре латеральной петли, слуховой тракт проходит через нижние холмики четверохолмия и медиальное коленчатое тело в первичную слуховую кору (поле 41 по Бродману). Таким образом, слуховой тракт состоит, по крайней мере, из пяти нейронов (рис. 14.5).
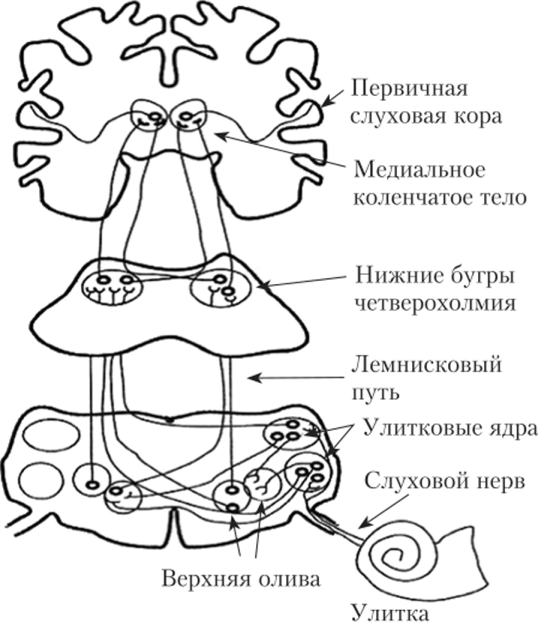
Рис. 14.5. Проводящие пути и переключательные ядра слуховой системы 1
Слуховой нерв содержит сравнительно мало волокон, но слух при этом остается острый. Это связано с двумя факторами:
1) каждое слуховое волокно, достигнув кохлеарного ядра в стволе мозга, делится на множество концевых ветвей, эти ветви строго тонотопически 1 [1]
распределяются в кохлеарном ядре (одна клетка имеет в кохлеарном ядре множественное представительство);
2) в разных частях кохлеарного ядра имеются разные типы синапсов и клеток. Главная часть содержит весьма крупные и характерные релейные нейроны, которые служат морфологическим субстратом разных видов переработки слуховой информации.
На уровне подкорковых центров слуха происходят процессы, обеспечивающие возможность пространственного (бинаурального) слуха. Для этого необходимо объединение информации, получаемой от правой и левой улиток. Первый уровень объединения информации от обеих улиток — комплекс верхней оливы. Он состоит из латерального (5-образного) и медиального (акцессорного) ядер. Функция нейронов латерального ядра состоит, в основном, в кодировании частотных характеристик звукового сигнала. Нейроны отвечают в основном на ипсилатсральную стимуляцию. Медиальное (акцессорное) ядро получает сигналы как от ипси-, так и от контрлатеральной улитки. Там есть нейроны-детекторы бинауральной временной задержки, которая отражает направление локализации источника звука.
Заднее двухолмие среднего мозга получает сигналы от дорсального кохлеарного ядра и латерального (акцессорного) ядра верхней оливы. Следовательно, механизмы пространственного слуха в двухолмии дублируют механизмы верхней оливы и обогащаются новыми детектирующими свойствами нейронов. Впервые здесь появляются нейроны, которые отвечают только при перемещении стимула (детекторы движения слухового сигнала).
Медиальное коленчатое тело таламуса (МКТ) тоже содержит много высоко селективных детекторов локализации и движения звука. Кроме того, звук у ряда нейронов МКТ помимо реакции на включение звука вызывает еще один компонент — реверберирующий разряд, который возникает через секунды после выключения звука и длится до нескольких секунд. Это связано с появляющимся только на этом уровне механизмом фиксации на достаточно длительное время информации о локализации звука в пространстве.
Анатомия и физиология дорсального кохлеарного ядра (ДКЯ)
дорсальное кохлеарное ядро (DCN, также известный как «tuberculum acusticum »), представляет собой корковидную структуру на дорсо-латеральной поверхности ствола мозга. Наряду с вентральным ядром улитки (VCN), оно образует ядро улитки (CN), где все волокна слухового нерва из улитки образуют свои первые синапсы.
Содержание
- 1 Анатомия
- 2 Физиология
- 3 Ссылки
- 4 Внешние ссылки
Анатомия
DCN отличается от вентральной части CN, поскольку она не только проецируется к центральному ядру (подразделению) нижнего бугорка (CIC), но также получает эфферентную иннервацию от слуховой коры, верхнего оливкового комплекса и нижнего colliculus. цитоархитектура и нейрохимия DCN аналогична таковой мозжечка, что является важным понятием в теориях функции DCN. Таким образом, считается, что DCN участвует в более сложной слуховой обработке, а не просто передает информацию.
пирамидные клетки или гигантские клетки являются основной группой клеток DCN. Эти ячейки являются целью двух разных систем ввода. Первая система возникает из слухового нерва и несет акустическую информацию. Второй набор входных сигналов передается через набор мелких гранулярных клеток в ядре улитки. Также имеется большое количество соседних ячеек колеса тележки. Гранулярные клетки, в свою очередь, являются мишенью для ряда различных входных сигналов, включая как те, которые участвуют в слуховой обработке, так и, по крайней мере, у низших млекопитающих, соматосенсорные входы, связанные с головой, ухом и челюстью.
Выступы основных клеток DCN образуют дорсальную акустическую полоску, которая в конечном итоге заканчивается в CIC. Эта проекция четко определенным образом перекрывается с проекцией боковой верхней оливы (LSO), где они формируют первичный возбуждающий вход для единиц ICC типа O.
Физиология
Основные ячейки в DCN имеют очень сложные кривые настройки интенсивности частоты. Классифицируемые как клетки кохлеарного ядра типа IV, частота возбуждения может быть очень высокой в ответ на звук низкой интенсивности на одной частоте, а затем упасть ниже спонтанной скорости с небольшим увеличением частота или интенсивность стимула. Затем частота стрельбы может увеличиваться с другим увеличением интенсивности или частоты. Ячейки типа IV возбуждаются широкополосным шумом и, в частности, возбуждаются шумовым стимулом непосредственно ниже наилучшей частоты ячейки (BF).
В то время как кустистые клетки VCN помогают локализовать звуковой стимул на горизонтальной оси через свои входы в верхний оливарный комплекс, клетки типа IV могут участвовать в локализации звукового стимула на вертикальная ось. ушная раковина избирательно усиливает частоты, что приводит к снижению звуковой энергии на определенных частотах в определенных областях пространства. Сложная схема срабатывания ячеек типа IV делает их особенно подходящими для обнаружения этих зазубрин, а с объединенной мощностью этих двух систем локализации обычный человек может определить место взрыва фейерверка без использования глаз.
Соматосенсорные входы подавляют активность клеток типа IV, возможно, подавляя их активность во время движений головы и ушной раковины. Хотя это не было широко изучено, это может играть важную роль в локализации источника звука на высоте. Аналогичный эффект наблюдается в зрительной системе в эффекте, известном как слепота к изменениям.
. Современные слуховые модели DCN используют модель с двумя ингибиторами. Клетки типа IV получают возбуждение непосредственно от слухового нерва и ингибируются клетками типа II (вертикальные) и широкополосным ингибитором (клетки начала-c).
Анатомия и физиология дорсального кохлеарного ядра (ДКЯ)
Анатомия и физиология дорсального кохлеарного ядра (ДКЯ)
Дорсальное кохлеарное ядро (ДКЯ) является центром мультисенсорной интеграции.37 Основные выходные нейроны биполярны и имеют булавовидные окончания на базальных и апикальных ветвях. Возбуждающие входы от улитки идут к базальным дендритам. Смешанные источники возбуждения и торможения связываются с апикальными дендритами и телами нейронов, поступая от вестибулярного, мозжечкового и соматосенсорного ядер.
Поскольку эти последние входы направляются к голове, шее и наружному уху, предполагается, что мозжечковая схема дорсального кохлеарного ядра (ДКЯ) может быть вовлечена в акустико-моторные поведенческие реакции и локализацию источника звука с основой в ушной раковине.
а) Анатомия дорсального кохлеарного ядра (ДКЯ). У большинства млекопитающих ДКЯ является трехслойной структурой. Наружный молекулярный слой содержит нисходящие входы, поступающих в ядро извне и местные целевые нейроны. Разнообразные источники этих «мшистых волокон» были определены при введении ПКХ в дорсальную акустическую полоску (ДАП), где волокна входят в ядро.
После этой процедуры, наблюдалось массивное ретроградное накапливание в комплексе верхней оливы, так как ПКХ подхватывался коллатеральными аксонами оливокохлеарного комплекса. Обширная маркировка неслуховых групп клеток была отмечена в мозжечке и вестибулярных ядрах. Ретроградное накапливание также было обнаружено в боковых клиновидных и сенсорных тройничных ядрах. В совокупности эти структуры несут проприоцептивную информацию от верхней части тела, лица и уха.
Мультисенсорная информация, доставленная к ДКЯ по мшистым волокнам, передается к клеткам-гранулам. Клетки-гранулы — это мелкие клетки, которые сосредоточены вдоль молекулярного слоя и в границах, которые разделяют главные части ядерного комплекса. Аксоны клеток-гранул формируют множество параллельных волокон, которые пересекают ядро на всем протяжении. Эти волокна возбуждают пирамидальные клетки посредством прямых связей с апикальными дендритами или тормозят их через местные интернейроны -колесообразные клетки.
Слой пирамидальных клеток образован крупными проецирующими нейронами ДКЯ. Пирамидальные клетки, известные как веретенообразные клетки, часто располагаются вдоль продольной оси ядра. Они посылают свои апикальные дендриты в скопление клеток молекулярного слоя, где с ними связываются клетки-гранулы и колесообразные клетки. Они направляют базальные дендриты в глубокий слой, где они встречают входы от слуховых нервных волокон и дополнительных источников торможения. Тонотопическая карта улитки передается к ДКЯ систематической иннервацией дорсальной порции слухового нерва.
Глубокий слой ДКЯ заполнен гигантскими и вертикальными клетками. Гигантские клетки — менее часто встречаемые проекционные нейроны. Основные физиологические свойства подобны пирамидальным клеткам, но их глубоко расположенные дендриты не достигают молекулярного слоя и не связываются с параллельными волокнами. Вертикальные клетки являются тормозящими интернейронами. Их узкополосное инг ибирующее влияние отражается расположением вдоль частотной полоски ДКЯ. Дополнительное широкополосное глицеринергическое торможение связано с лучистыми клетками вентрального кохлеарного ядра (ВКЯ).

Анатомическое распределение первичных морфологических классов в рамках слоистой структуры дорсального кохлеарного ядра.
С — веретенообразная клетка; G — клетка-гранула; Р — пирамидная клетка; pf — параллельные волокна;
PVCN — задневентральное кохлеарное ядро; R — лучистая клетка; V — вертикальная клетка.
б) Нейронное кодирование спектральных сигналов для звуковой локализации. Основные физиологические свойства проекционных нейронов дорсального кохлеарного ядра (ДКЯ) формируются сложными взаимодействиями их многочисленных возбуждающих и тормозящих входных сигналов. Изображения ВКЯ линейные, узко настроенные и рассредоточенные, изображения ДКЯ нелинейные, широко настроенные и интегративные. Поэтому показатели разряда индивидуальных пирамидальных и гигантских клеток идеально подходят для кодирования спектральной формы сложных звуков.
Кошка, являющаяся наиболее изученной животной моделью обработки в дорсальном кохлеарном ядре (ДКЯ), соответствующую спектральную информацию получает посредством передаточной функции слухового аппарата (ПФСА).
Эта форма фильтра описывает направленные ответы головы и наружного уха на свободные стимулы, при их распространении к барабанной перепонке. Основанные на ПФСА спектральные сигналы имеют решающее значение для точной локализации высоты источника звука.
Характерной особенностью ПФСА кошки является единственный глубокий спектральный зубец, который резко уменьшается на частотах между 5 и 20 кГц. Когда широкополосной звук проходит через передаточную функцию, эти частоты по существу, отфильтровываются из стимула. Частотная локализация зубца систематически меняется при движении источника звука по высоте или по азимуту. Когда эти сигналы удаляются из свободного звукового поля, то кошки демонстрируют слабонаправленные ориентационные ответы.
Нейронное кодирование спектральных сигналов для локализации звука было исследовано при записи ПФСА-у-правляемых показателей разряда проекционных нейронов дорсального кохлеарного ядра (ДКЯ), которые известны как IV тип нейронов. Для улучшения контроля стимула эти эксперименты с моделируемыми спектральными зубцами выполнялись с помощью наушников. Зубец может быть смещен по частоте методами обработки цифрового сигнала в соответствии с настройкой нейронов, расположенных вдоль траектории регистрирующего электрода.
Целью этих экспериментов являлась имитация перемещения источника звука, путем манипулирования функции интересов по отношению к ингибирующим свойствам IV типа нейронов. Когда частота зубца ниже оптимальной частоты (ОЧ), нейрон стимулируется исключительно от раздражителя. Аналогичный ответ получают, когда зубец выше ОЧ . Такие результаты ожидаются при учете возбуждающего эффекта широкополосного шума. Однако, когда зубец совпадает с ОЧ, то нейрон в значительной степени ингибируется.
в) Симптомы и клиника поражения дорсального кохлеарного ядра (ДКЯ). Перцептивные нарушения, связанные с нарушениями обработки в ДКЯ, были исследованы в лаборатории путем пересечения дорсального кохлеарного тракта (ДКТ). Эта хирургическая манипуляция устраняет восходящую аксональную инневрацию пирамидальными и гигантскими клетками нижнего бугорка. В отличие от поражений трапециевидного тела, поражения ДКТ мало влияют на основные признаки слуха. Абсолютные пороги в тишине и на фоне шума не зависят от данной процедуры.
Значительные нарушения наблюдаются в тот момент, когда кошек с поражением дорсального кохлеарного тракта (ДКТ) тестируют на локализацию парадигм, требующих идентификации высоты источника звука. Кошки делают рефлекторные движения головой по направлению к внезапным, неожиданным звукам. Они также могут быть обучены согласованному выполнению данных движений при поощрении пищей. Кошка хищник по своей природе, и точность этих управляемых звуком реакций приближается к границам человеческого направленного слуха.
Когда выходные проводящие пути дорсального кохлеарного ядра (ДКЯ) с двух сторон разрушены, кошки демонстрируют явные проблемы с ориентацией. Несмотря на то, что выполнение находится под угрозой и в горизонтальном, и в вертикальном направлениях, наибольшие ошибки наблюдаются в определении высоты источника звука. Эти систематические ошибки подтверждают, что ДКЯ является важным участком для слуховой обработки спектральных сигналов с целью определения локализации звука.
Кошки с двусторонними поражениями дорсального кохлеарного тракта (ДКТ) сохраняют направленный слух в горизонтальном направлении путем участия бинауральных сигналов, которые обрабатываются в вентральных путях, ведущих к комплексу верхней оливы.
У людей редко встречается полное торможение выходов ДКЯ при травме сосудов. Тиннитус (шум в ушах) может отражать не полностью картину разрушения этой деликатной структуры. Если текущие физиологические обоснования шума в ушах подтвердятся, то нарушения обработки в ДКЯ отражают наиболее частую форму нарушений слуха в современном индустриальном обществе.

Эффекты от повреждения дорсального кохлеарного тракта (ДКТ) на слуховую ориентацию.
А — место поражения. Б — точность слуховой ориентации у интактной кошки.
В — Точность после повреждения дорсального кохлеарного тракта (ДКТ) с обеих сторон.
Эллипсы указывают стандартное отклонение реакций в горизонтальных и вертикальных измерениях.
Существует достаточно доказательств того, что частота спонтанных разрядов в ДКЯ увеличивается, когда кохлеарные входы уменьшаются вследствие кратковременного воздействия интенсивного звука или долгосрочных процессов старения. Поскольку эта гиперактивность имитирует нормальную слуховую реакцию на физический стимул, это создает впечатление фантомного звука.
Функциональные изменения, сопровождающие звон в ушах, были тщательно изучены на моделях животных, позволяя контролировать индукцию расстройства и непосредственную физиологическую оценку последствий. Этот анализ пока не раскрывает однозначный участок источника и редкую патологию, но становится ясно, что потенциальные структуры имеют общие свойства. ДКЯ, НК и слуховая кора связаны с тиннитусом.
Эти центры обработки являются ключевыми участками мультисенсорной интеграции, где входы от слуховых и неслуховых ядер сходятся для того, чтобы действовать в совокупности возбуждающих и тормозных нейромедиаторов. Нарушения любого компонента в этой синаптической конструкции способны перестроить состояние покоя и звукопроводящие свойства нейронной цепи.
Нейронная гиперактивность остается определяющей физиологической моделью тиннитуса. Широкая поддержка этого гипотетического механизма отчасти связана с интуитивно понятной простотой. Кроме того, глобальные изменения в мозговой активности предсказывают метаболическую причину, которая может быть обнаружена неинвазивно у пациентов с шумом в ушах.
Несмотря на то, что некоторые модели на животных реагируют на акустическое чрезмерное воздействие при помощи генерализованной гиперактивности, важные детали предложенной связи между активностью головного мозга и восприятием звука остаются неразрешенными. Результаты могут быть специфичными для видов, способов индукции и методов электрофизиологической записи. Более полное объяснение основных механизмов звона в ушах может быть получено из более сложного статистического анализа моделей регулярности, синхронизации, либо долгосрочных колебаний спонтанной активности. Эти свойства не могут быть адекватно описаны без прямых электрофизиологических записей на моделях животных.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Дорсальное кохлеарное ядро - Dorsal cochlear nucleus
Дорсальное ядро кохлеарного ( DCN , также известное как « бугорок acusticum »), представляет собой кортекс-подобную структуре на дорсо-боковой поверхности ствола головного мозга. Наряду с вентральным ядром улитки (VCN) он образует ядро улитки (CN), где все волокна слухового нерва из улитки образуют свои первые синапсы.
СОДЕРЖАНИЕ
Анатомия
DCN отличается от вентральной части CN, поскольку он не только проецируется в центральное ядро (подразделение) нижнего бугорка (CIC), но также получает эфферентную иннервацию от слуховой коры , верхнего оливкового комплекса и нижнего бугорка. Цитоархитектура и нейрохимия из DCN подобна тому из мозжечка , важного понятия в теории функции DCN. Таким образом, считается, что DCN участвует в более сложной слуховой обработке, а не просто в передаче информации.
В пирамидальных клетках или гигантские клетки являются основной ячейкой группировки DCN. Эти ячейки являются целью двух разных систем ввода. Первая система возникает из слухового нерва и несет акустическую информацию. Второй набор входных сигналов передается через набор мелких гранулярных клеток в ядре улитки. Есть также множество соседних ячеек с колесами . Гранулярные клетки, в свою очередь, являются мишенью для ряда различных входных сигналов, включая как те, которые участвуют в слуховой обработке, так и, по крайней мере, у низших млекопитающих, соматосенсорные входы, связанные с головой, ухом и челюстью.
Проекции от основных клеток DCN образуют дорсальную акустическую полоску, которая в конечном итоге заканчивается в CIC. Эта проекция четко определенным образом накладывается на проекцию боковой верхней оливы (LSO), где они формируют первичный возбуждающий вход для единиц ICC типа O.
Физиология
Основные ячейки в DCN имеют очень сложные кривые настройки интенсивности частоты. Классифицируемые как клетки ядра улитки IV типа, частота возбуждения может быть очень высокой в ответ на звук низкой интенсивности на одной частоте, а затем падать ниже спонтанной частоты только с небольшим увеличением частоты или интенсивности стимула. Затем частота стрельбы может увеличиваться с другим приращением интенсивности или частоты. Клетки типа IV возбуждаются широкополосным шумом и, в частности, возбуждаются шумовым стимулом непосредственно ниже наилучшей частоты (BF) клетки.
В то время как кустистые клетки VCN помогают локализовать звуковой стимул на горизонтальной оси через свои входы в верхний оливарный комплекс , клетки типа IV могут участвовать в локализации звукового стимула на вертикальной оси. Ушная раковина селективно усиливает частоты, что приводит к снижению звуковой энергии на определенных частотах в определенных областях пространства. Сложные схемы срабатывания ячеек типа IV делают их особенно подходящими для обнаружения этих зазубрин, а с объединенной мощностью этих двух систем локализации обычный человек может определить место взрыва фейерверка, не используя своих глаз.
Соматосенсорные входы подавляют активность клеток типа IV, возможно, подавляя их активность во время движений головы и ушной раковины. Хотя это не было широко изучено, это может играть важную роль в локализации источника звука на высоте. Аналогичный эффект наблюдается в зрительной системе в эффекте, известном как слепота к изменениям .
Современные слуховые модели DCN используют модель с двумя ингибиторами. Клетки типа IV получают возбуждение непосредственно от слухового нерва и ингибируются клетками типа II (вертикальные) и широкополосным ингибитором (клетки начала-c).
3.3. Центральный отдел слуховой системы
В работах С. Рамон-и-Кахаля дано описание классического слухового пути. Этот восходящий специфический (или афферентный) путь представляет собой несколько последовательных уровни (рис. 22):
1. спиральный ганглий улитки;
2. кохлеарные ядра продолговатого мозга;
3. верхняя олива продолговатого мозга;
4. нижние бугры четверохолмия среднего мозга;
5. медиальные коленчатые тела таламуса;
6. слуховые поля височной коры головного мозга.
Кроме классического пути были обнаружены дополнительные водящие слуховые пути, связанные с ретикулярной формацией ствола, мозжечком, ассоциативными ядрами таламуса и корковыми полями, окружающими центральное слуховое поле. Наряду с восходящими путями существует также система нисходящих слуховых путей, идущих от слуховой коры к нижележащим слуховым

образованиям. Слуховая система имеет многочисленные связи с другими сенсорными системами и моторными центрами.
Спиральный ганглий улитки и кохлеарный нерв. Первый уровень в восходящем слуховом пути составляют клетки спирального ганглия улитки, являющиеся нейронами I порядка. Эти биполярные (с двумя отростками) клетки, расположенные в виде тяжа плотно упакованных компактных групп, находятся в спиральном канале улитки параллельно органу Корти. Спиральный ганглий по длине значительно меньше, чем орган Корти, и делает меньше оборотов вокруг оси улитки. Тела всех клеток спирального ганглия и их отростки покрыты миелиновыми оболочками (как телефонный кабель).
Периферические отростки клеток спирального ганглия направляются к органу Корти, а центральные выходят во внутренний слуховой проход и образуют кохлеарный (или слуховой) нерв.
Кохлеарные ядра продолговатого мозга. В них переключаются все волокна слухового нерва. Он представляет собой слуховую ветвь VIII пары черепно-мозговых нервов. Слуховой нерв входит в продолговатый мозг и оканчивается на клетках кохлеарных ядер. В кохлеарном комплексе выделяют три основные ядра различной структуры: 1. переднее вентральное; 2. заднее вентральное; 3. дорсальное (или слуховой бугорок).
Ядра представляют собой скопление нейронов II порядка. Волокна слухового нерва при входе в кохлеарные ядра разделяются на две ветви — нисходящую и восходящую, иннервирующие соответственно клетки заднего и переднего ядер. Волокна от апикальной части улитки иннервируют вентролатеральную часть кохлеарного комплекса, а от базальной части — его дорсомедиальную область.
Дорсальное ядро кохлеарного комплекса по структуре резко отличается от вентральных ядер слоистым строением, сходным с корой, и сложным характером внутренних связей, а также обилием разветвлений нисходящих волокон. В кохлеарных ядрах находится большое число ассоциативных клеток.
От клеток кохлеарных ядер начинаются контрлатеральный и ипсилатеральный восходящие слуховые пути, но основная масса волокон переходит на противоположную сторону (рис. 23).
Контрлатеральный путь образует три пучка волокон, образующих слуховые полоски:
1. вентральную (трапециевидное тело); 2. интермедиальную (полоска Хельда); 3. дорсальную (полоска Монакова).
Основную часть проекционных волокон содержит трапециевидное тело. Волокна оканчиваются в различных слуховых образованиях: основная часть — на клетках верхней оливы, другие — на ядрах латеральной петли, небольшая часть - в нижних буграх четверохолмия и медиальном коленчатом теле.

Верхняя олива. В комплекс ядер верхней оливы входят: 1) S-образное (латеральное) ядро; 2) медиальное (акцессорное) ядро; 3) группа периоливарных ядер.
S-образное ядро получает афферентацию только от ипсилатерального кохлеарного ядра, в то время как аксоны его клеток иннервируют выше лежащие ядра как своей, так и противоположной стороны.
Клетки акцессорного ядра имеют два дендрита; на медиальном дендрите оканчиваются аксоны клеток переднего кохлиарного ядра противоположной стороны, а на латеральном дендрите — аксоны клеток того же ядра той же стороны. Таким образом, акцессорное ядро верхней оливы является первым уровнем слуховой системы, осуществляется бинауральная конвергенция афферентации от правой и левой улиток.
Группа периоливарных ядер (6 — 7 ядер) получает связи от переднего заднего вентральных кохлеарных ядер своей и противоположной сторон. Аксоны клеток этих ядер направляются в основан на свою сторону.
Волокна дорсальной и интерлатеральной полосок, а также аксоны клеток оливы и непереключенные волокна трапециевидного тела образуют мощный пучок — латеральную петлю.
Латеральная петля направляется вверх от оливы и подходит к нижним буграм четверохолмия. Часть волокон латеральной петли
оканчивается на клетках заднего бугра, а другая — в таламусе. Небольшое число волокон латеральной петли направляется на противоположную сторону, образуя комиссуру Пробста. Латеральная петля представляет собой общий путь для аксонов нейронов II, III и отчасти IV порядков и представляет улитку как своей, так и противоположной стороны. Латеральная петля — основной коллектор аксонов слуховых ядер продолговатого мозга.
Нижние бугры четверохолмия — среднемозговой центр слуховой системы (иначе называется задним, или нижним, двухолмием) состоит из нейронов III, IV, V порядков. Структура этого образования очень сложная. Основная часть клеточных элементов заднего холма расположена в центре и составляет его центральное ядро. Аксоны клеток заднего холма направляются к медиальному коленчатому телу своей стороны и частично к противоположному заднему холму.
Медиальные коленчатые тела — таламический центр слуховой системы. В нем различают крупно- и мелкоклеточную (главную) части. Мелкоклеточная часть, через которую проходит основной слуховой путь, неоднородная и подразделяется на дорсальную, вентральную и медиальную области. Для дорсальной области характерны крупные нейроны с толстыми дендритами, отходящими от тела клетки в различных направлениях и образующими сферическое дендритное поле. Для вентральной области основными являются нейроны с пучковидными дендритами, которые отходят от тела клетки только в одной плоскости, образуя дендритное поле дисковидной формы.
В медиальной области встречаются нейроны обоих типов, а также нейроны промежуточной формы.
Вентральная область имеет наибольшие размеры, и в ней оканчивается большая часть входящих волокон. Здесь была обнаружена ламинарная структура (область синаптических контактов), причем клетки каждого слоя иннервируются афферентными волокнами с двух сторон. Аксоны клеток медиального коленчатого тела образуют акустическую радиацию и оканчиваются на клетках слуховой коры. Комиссура Гуддена связывает оба коленчатые тела.
Итак, путь электрических импульсов от рецепторов органа слуха к слуховой коре больших полушарий головного мозга имеет 3 — 5 уровней переключения (переключательных станций) и не менее трех перекрестов (рис. 24).
Информация о звуковом потоке, попадающем в диапазон возможностей рецепторной части органа слуха по аксонам нервных клеток слухового спирального ганглия, дендриты которых подходят к рецепторным клеткам, передается в слуховой центр продолговатого мозга (кохлеарные ядра) в форме коротких электрических импульсов. Они распространяются вдоль волокон слухового нерва со скоростью от 0,5 до 100 м/с — в зависимости от толщины волокон.

После переключения волокон слухового нерва на клетках кохлеарных ядер продолговатого мозга электрические импульсы поступают к следующему клеточному скоплению – ядрам верхней оливы. Здесь отмечается первый перекрёст слуховых путей: меньшая часть волокон остаётся в пределах полушария, на стороне которого расположены периферические слуховые рецепторы, а большая часть идет в противоположное полушария головного мозга. В области основания мозга, где располагается перекрест, имеется еще одна группа ядер — ядра трапециевидного тела. В них также осуществляется частичное переключение волокон клеток кохлеарных ядер. Небольшая часть этих волокон направляется, переключаясь, в средний мозг и заканчивается на клетках нижних (задних) холмов. Сюда же приходит значительная часть перекрещенных и не перекрещенных волокон из ядер верхней оливы.
Подавляющее большинство волокон от клеток кохлеарных ядер, верхнеоливарных ядер, трапециевидного тела и латеральной петли переключается на клетках нижних холмов, после чего волокна следующего порядка либо переходят в противоположное полушарие (второй крупный перекрест), либо идут непосредственно к ближайшим подкорковым слуховым центрам таламуса (зрительного бугра) — медиальным коленчатым телам. Только очень небольшая волокон проходит мимо нижних холмов, не переключаясь в них, и заканчивается прямо в медиальном коленчатом теле.
Читайте также: